

دوره 30، شماره 12 - ( اسفند 1401 )
جلد 30 شماره 12 صفحات 6210-6199 |
برگشت به فهرست نسخه ها


Ethics code: IR.IAU.FALA.REC.1400.023
![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Ahmadzadeh Chaleshtori Z, Rastegari A A, Nayeri H, Doosti A. Evaluation of the Cytokine Genes Expression in Vaccinated BALB/c Mice with pEGFP-C2-leoA DNA Vaccine. JSSU 2023; 30 (12) :6199-6210
URL: http://jssu.ssu.ac.ir/article-1-5762-fa.html
URL: http://jssu.ssu.ac.ir/article-1-5762-fa.html
احمدزاده چالشتری زهرا، رستگاری علی اصغر، نیری هاشم، دوستی عباس. بررسی تغییرات بیان سایتوکاینها در موشهای BALB/c واکسینه شده با واکسن ژنی pEGFP-C2-leoA. مجله علمي پژوهشي دانشگاه علوم پزشكي شهید صدوقی يزد. 1401; 30 (12) :6199-6210



متن کامل [PDF 1132 kb]
(564 دریافت)
| چکیده (HTML) (1545 مشاهده)
متن کامل: (1061 مشاهده)
مقدمه
هلیکوباکتر پیلوری (H. pylori) یکی از شایعترین پاتوژنهای انسانی است زیرا مخاط معده حدود 50 درصد از جمعیت جهان را آلوده میکند (1). اکثر عفونتهای ایجاد شده بدون علامت هستند و بیشتر در جمعیت اطفال ایجاد میشوند که عفونت را بدون ریشهکنی مؤثر باکتریایی مادامالعمر میسازد. علاوه بر این، مطالعات اپیدمیولوژیک که عفونت را با خطر بالاتر بدخیمی معده مرتبط میکند، سازمان بهداشت جهانی را برای تحقیقات سرطان جهت طبقهبندی هلیکوباکتر پیلوری بهعنوان سرطانزای کلاس I هدایت میکند (2). عفونت هلیکوباکتر پیلوری یک عامل خطر اصلی برای توسعه سرطان معده است زیرا باعث ایجاد یک توالی گامبهگام در مخاط معده میشود که با گاستریت سطحی شروع میشود، که میتواند به گاستریت مزمن، گاستریت آتروفیک، متاپلازی روده، دیسپلازی و درنهایت کارسینوم معده تبدیل شود (4،3). باکتریها پاسخ ایمنی میزبان (ذاتی و سازگار) را القا میکنند، اما تداوم عفونت نشان میدهد که پاسخ در از بین بردن عفونت مؤثر نیست. علاوه بر این، شواهد متعدد نشان میدهد که پاسخ ایمنی به پاتوژنز مرتبط با عفونت کمک میکند. در طول دو دهه گذشته، چندین مدل تجربی از عفونت هلیکوباکتر پیلوری برای بررسی پاتوژنز این عفونت ایجادشده است. با استفاده از سویه هلیکوباکتر پیلوری سازگار با موش (Sidney Strain،SS1)، لی و همکاران. مدلی از کلونیزاسیون باکتریایی طولانیمدت و بالا را در موشها ایجاد کردند (3). اگرچه هلیکوباکتر پیلوری بهعنوان سویه غیرتهاجمی شناخته شده است، اما باعث ایجاد یک واکنش التهابی گسترده در مخاط معده میشود (5). این واکنش با نفوذ مخاطی سلولهای التهابی، بهویژه نوتروفیلها، مشخص میشود که با افزایش بیان کموکاینها و سیتوکینهای پیش التهابی انجام میشود (7،6). با توجه به شیوع بالای عفونت هلیکوباکتر پیلوری در سراسر جهان، هزینههای بالای درمان آنتیبیوتیکی و افزایش نرخ مقاومت آنتیبیوتیکی، تلاشهای قابلتوجهی برای تولید واکسن علیه هلیکوباکتر پیلوری انجام شده است. بااینحال، واکسیناسیون کارآمد تا به امروز در انسان به دست نیامده است (8). بنابراین، یافتن درمانهای جایگزین ضروری است (9). ژن leoA یکی از عوامل بیماریزایی GTPase هلیکوباکترپیلوری است که در جزیره بیماریزایی کدگذاری میشود و بالقوه از طریق وزیکول ترشحی باعـث افـزایش انتشـار سـم مـیشـود (10). وزیکولهای غشاء نقش مهمـی در عملکـرد حفاظتشده اعضـای خانواده دیآمینی، دارند. گزارشات نشان دادهاند که این ژن توانایی ایمنیزایی بالقوه در بدن میزبان را دارد؛ لذا آن را نامزد مناسبی برای تولید یک واکسن ژنی علیه هلیکوباکترپیلوری، معرفی نمودهاند. هدف از این مطالعه بررسی میزان بیان ژنهای IL6، IL4 و اینترفرون گاما در سطح mRNA به روش real time PCR در خون موشهای BALB/c واکسینه شده با واکسن ژنی pEGFP-C2-leoA بوده است. همچنین میزان بیان و ماندگاری leoA در بافت محل تزریق واکسن (عضله ران موشها) در یک دوره زمانی ۴۵ روزه بررسیشده است.
روش بررسی
کلونسازی ژن در وکتور بیانی: در این مطالعه، قطعه ژن leoA هلیکوباکترپیلوری درون وکتور بیانی pEGFP-C2 طراحی و توسط شرکت Generay سنتز گردید. باکتریE. coli سویه Top10F بهمنظور ترانسفورماسیون و تکثیر سازههای ژنی نوترکیب مورداستفاده قرار گرفت. انتخاب و غربالگری باکتریهای دریافتکننده پلاسمید نوترکیب در حضور آنتیبیوتیک کانامایسین انجام شد. صحت وکتور نوترکیب (pEGFP-C2-leoA) به روش PCR و هضم آنزیمی با آنزیمهای KpnI و SacII تأیید گردید. همچنین صحت توالی ژن کلون شده درون وکتور بیانی pEGFP-C2، توسط شرکت سازنده و با روش تعیین توالی سانگر موردبررسی قرار گرفت.
انتقال سازواره نهایی pEGFP-C2-leoA به سلولهای جانوری: بهمنظور بررسی بیان ژن leoA در سلول جانوری، از سلول CHO استفاده شد و برای ترانسفورمیشن این سلولها از روش الکتروپوریشن (مدل Gene Pulser Xcell ساخت شرکت Bio Rad) بهره گرفته شد. تعداد دو میلیون سلولهای CHO به همراه ۸۰۰ نانوگرم در هر میکرو لیتر از وکتور نوترکیب pEGFP-C2-leoA و وکتور pEGFP-C2 بدون ژن در دو گروه مختلف در حجم ۴۰۰ میکرولیتر در کوت 0/4 میلیمتری ریخته شد. پالس الکتریکی با شرایط بهینهسازی شده 0/174 کیلوولت و ۴۰۰ میکرو فاراد به سلولها داده شد و سلولهای حاصل در حضور آنتیبیوتیک نئومایسین کشت داده شدند.
انجام SDS-PAGE: وزن مولکولی و محل قرارگرفته پروتئین بیانشده درون سلولهای یوکاریوتی CHO، بهوسیله آزمون SDS-PAGE تأیید شد. سلولهای CHO ترانسفورم شده به مدت ۳ دقیقه با دور ۳۰۰۰ دور بر دقیقه رسوب داده شدند. به رسوب سلولی مقدار ۱۰۰ میکرو لیتر PBS اضافه شد و در نهایت سلولهای حاصل بر روی ژل ۱۲ درصد SDS-PAGE الکتروفورز و با کوماسی بلو رنگآمیزی انجام گرفت.
تهیه نانو ذرات کیتوزان به روش Ionic Gelation: ابتدا محلول کیتوزان (حلکردن صد میلیگرم کیتوزان در ۵۰ میلیلیتر اسید استیک به مدت ۲۴ ساعت در دمای اتاق با ۵/۵= pH و فیلتر 45/0 میکرومتری جهت حذف کیتوزانهای حلنشده) تهیه شد. سپس محلول TPP (۲۰میلیگرم در ۲۰ میلیلیتر آب دیونیزه) بهصورت قطرهقطره به محلول کیتوزان در حال چرخش روی همزن در مدت ۱ ساعت با دور rpm ۱۰۰۰ اضافه شد. سپس محلول حاصل به مدت ۱۵ دقیقه با دور rpm ۱۴۰۰۰سانتریفیوژ شد و محلول رویی حاوی نانو ذرات کیتوزان جمعآوری گردید و با دستگاه فریز - درایر خشک و پودر گردید. از روشهای زتا آنالایزر (دستگاه Nanozeta Sizer ساخت کشور آمریکا) برای ارزیابی ویژگی-های فیزیکوشیمیایی، شاخص پراکندگی و اندازه نانو ذرات کیتوزان تولیدشده استفاده شد و از دستگاه Malvern Instruments ساخت کشور انگلستان و میکروسکوپ الکترونی (SEM) برای پراکندگی نوری دینامیک (Dynamic Light Scattering) استفاده گردید. تهیه مخلوط پلاسمید و نانو ذرات: نسبت مساوی از محلول کیتوزان (۱ درصد) و پلاسمید (۲۰۰۰ میکروگرم در هر میلیلیتر PBS) باهم مخلوط شدند و به مدت ۱ ساعت در دمای ۵۵ درجه سانتیگراد قرار گرفتند.
گروهبندی و زمانبندی تزریق به موشها
18 سر موشهای ماده BALB/c ششهفتهای با وزن 30-22 گرم به سه گروه ۶تایی دستهبندی شدند:
- گروه اول: پلاسمید نوترکیب (واکسن ژنی) + نانو ذرات کیتوزان
- گروه دوم: پلاسمید نوترکیب (واکسن ژنی)
- گروه سوم: پلاسمید فاقد ژن هدف (گروه شاهد)
تزریق گردید. تزریقها بافاصله زمانی ۰، ۷ و ۱۵ روز انجام گرفت.
تزریق به موش: موشهای گروه اول مقدار ۱۰۰ میکرولیتر (حاوی ۱۰۰ میکروگرم DNA)، و موشهای گروه دوم ۱۰۰ میکرولیتر (حاوی ۱۰۰۰ میکروگرم در هر میلیلیتر PBS را بهصورت تزریقی دریافت کردند. به گروه شاهد نیز محلولی بهصورت ۱۰۰۰ میکروگرم در هر میلیلیتر PBS تهیه شد. سپس ۱۰۰ میکرولیتر از آن تزریق گردید.
نمونهبرداری: در روزهای پانزدهم، سیام و چهل و پنجم پس از آخرین مرحله تزریق، تعداد ۲ سر موش از هر گروه، کشته و نمونهبرداری شد.
بافت عضله ران موش) دقیقاً در محل تزریق(، و مونوسیتهای خون کامل با استفاده از فایکول جداسازی شد.
استخراج RNA: با استفاده از کیت استخراج RNA (شرکت یکتا تجهیز)، RNA کلی بافت طبق دستورالعمل شرکت سازنده تخلیص شد و آن با استفاده از الکتروفورز روی ژل آگارز ۱ درصد و مشاهده باندهای 18S و 28S بررسی گردید. همچنین غلظت RNAها با استفاده از دستگاه نانودراپ سنجیده شد. بلافاصله جهت سنتز cDNA مورداستفاده قرار گرفت.
ساخت Cdna: برای سنتزcDNA ، یک میکروگـرم از RNA استخراجشده با اسـتفاده از آغـازگر الیگـومر تیمیدین (Oligo dT) و کیـت شــرکت تاکارا ژاپن، در حجم ۱۰ میکرولیتر طبـق دستورالعمل شــرکت ســازنده بــه cDNA تبدیل گردید.
واکنش RT-qPCR: پرایمرهای مورداستفاده توسط نرمافزار Oligo 7 و برنامه Blast طراحی شدند (جدول ۱). هر واکـنش PCR (شامل SYBR Green Master Mix، پرایمر، cDNA و آب مقطر) در ۴۵ سیکل انجام شد و از ژن GAPDH بهعنوان ژن رفرنس در تمام آزمایشات بیان ژن استفاده شد.
تجزیهوتحلیل آماری
نتایج بهدستآمده بهوسیله نرمافزار SPSS version 16 مورد تحلیل آماری قرارگرفت؛ از آزمون one way ANOVAو آزمون متعاقب LSD و نیز آزمونمستقل t-test ، جهت بررسی و وجود ارتباط و میزان معنیداری دادهها استفاده گردید. تمام دادهها بهصورت means + S.E.M در سطح معنیداری p<0.05 در نظر گرفته شدند. آنالیز دادهها با استفاده از متد CtΔΔ-۲ انجام شد. مزیت روش ΔΔCT در سهولت مراحل کار است و یکبار رسم منحنی استاندارد کافی میباشد و در صورتی از این روش استفاده میشود که کارایی PCR در نمونههای کنترل و هدف نزدیک به 100٪ باشد.
ملاحظات اخلاقی
مطالعه حاضر از نوع تجربی میباشد که در کمیته اخلاق دانشگاه آزاد اسلامی واحد فلاورجان با کدIR.IAU.FALA.REC.1400.023 به تصویب رسیده است.
جدول 1: توالی پرایمر استفادهشده در این تحقیق

نتایج
صحت کلون شدن ژن leoA: درستی سازواره طراحیشده با روشهای PCR و هضم آنزیمی (KpnI و SacII) مورد تأیید قرار گرفت. برش وکتور pEGFP-C2-leoA با دو آنزیم فوقالذکر سبب تشکیل دو قطعه، با اندازههای ۶۴۹۲ جفت باز و ۱۷۵۷ جفت باز گردید که نشاندهنده تشکیل سازواره نهایی بود (شکل ۱).
الکتروپوریشن و بیان ژن: درستی ورود سازواره pEGFP-C2-leoA در سلولهای CHO با واکنش PCR و رشد در حضور آنتیبیوتیک تأیید شد. بیان ژن leoA در این سلولها، با واکنش SDS-PAGE انجام شد. محصول پروتئینی حاصل از ژن leoA به وزن ۱۲۴ کیلو دالتون نشاندهنده بیان این پروتئین در سلولهای یوکاریوتی CHO بود (شکل ۱).
تأیید نانو ذرات کیتوزان: اندازه نانو ذرات کیتوزان (وزن مولکولی پایین) با استفاده از میکروسکوپ الکترونی کمتر از ۲۰۰ نانومتر به دست آمد و میانگین اندازه نانو ذرات بهدست آمده توسط پراکنگی نوری دینامیک (DLS) 111/7 نانومتر و همچنین شاخص پراکنگی اندازه ذرات توسط آنالیز زتا (mV20/8) تعیین شد.
بررسی بیان ژنها: ژن leoA: بیان این ژن در نمونه بافت بهدستآمده از عضله ران موشهای واکسینه شده به روشreal time RT-PCR انجام شد. منحنیهای ذوب مربوط به ژنهای leoA و GAPDH بهصورت تک قله به دست آمد که بیانگر وجود تنهـا یـک محـصول خاص در PCR است. مقایسه بیان ژن leoA در بافت موشهای گروه حاوی پلاسمید با گروه حاوی نانو ذرات کیتوزان در روز ۱۵ افزایش بیان داشت و در روز ۴۵ کاهش بیان را نشان داد (نمودار ۱).
بیان ژنهای IL6، γIFN و IL4: بیان این ژنها در نمونه خون موشهای واکسینه شده با روش real time RT-PCR اندازهگیری شد. منحنیهای ذوب مربوط به ژنهای IL6، IFNᵧ، و IL4بهصورت تک قله به دست آمد که بیانگر وجود یـک محـصول خاص درPCR است. مقایسه میانگین بیان نسبی این ژنها نشان داد IFNᵧ در نمونههای پلاسمید و نمونه پلاسمید به همراه نانو ذرات کیتوزان نسبت به نرمال، افزایش بیان دارد. همچنین IL6 در نمونههای پلاسمید و نمونههای پلاسمید به همراه نانو ذرات کیتوزان نسبت به نرمال، کاهش بیان نسبت به نرمال نشان داد (نمودار ۱).
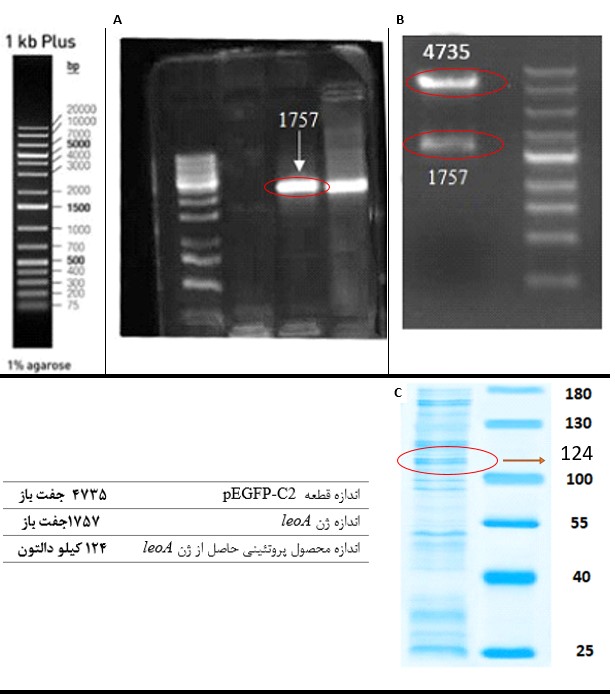
شکل 1: A) تکثیر ژن leoA به روش PCR، B) هضم آنزیمی سازواره نهایی pEGFP-C2-leoA با دو آنزیم KpnI و SacII و C) محصول پروتئینی حاصل از ژن leoA


نمودار 1: A: بررسی بیان ژن leoA در بافت موشهای حاوی پلاسمید همراه با ژن که در زمانهای 15، 30 و 45 بهصورت معنیدار (P <.0.01) اختلاف دارند. B) بررسی بیان ژن leoA در بافت موشهای حاوی پلاسمید همراه با ژن همراه با نانوذره کیتوزان که در زمانهای 15، 30 و 45 بهصورت معنیدار (P<.0.01) اختلاف دارند. C) بررسی بیان سایتوکاینها در مونوسیت استخراجشده از خون کامل موشهای حاوی پلاسمید همراه با ژن همراه با نانوذره که در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IFNᵧ بهصورت معنیدار افزایشیافته است (P<.0.01). در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IL6 بهصورت معنیدار افزایشیافته است (P<.0.5). در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IL4 بهصورت معنیدار کاهشیافته است (P<.0.01). D) بررسی بیان سایتوکاینها مونوسیت استخراجشده از خون کامل موشهای حاوی پلاسمید همراه با ژن که در گروه pEGFP-C2-leoA بیان ژن IFNᵧ بهصورت معنیدار (P<.001) افزایشیافته است، در گروه pEGFP-C2-leoA بیان ژن IL6 بهصورت معنیدار (P<0.5) تغییر نداشته است و در گروه pEGFP-C2-leoA بیان ژن IL4 بهصورت معنیدار (P<.0.01) کاهشیافته است.
بحث
هلیکوباکترپیلوری یک باکتری گرم منفی، متحرک، میلهای شکل و میکروآئروفیل (باکتریهایی که اکسیژن را بهعنوان گیرنده نهایی الکترون نیاز دارند و در شرایط بیهوازی به مقدار خیلی کمی رشد میکنند) است، که در سطح مجرایی پوشش معده دیده میشود (11). این باکتری دلیل اصلی گاستریت فعال و مزمن، زخم معده و دوازدهه است. همچنین موجب التهاب طولانی در مخاط معده بهوسیله نوتروفیلها، لنفوسیتها و پلاسماسلها میشود (12). ازآنجاییکه این باکتری در لایه عمقی موکوس پوشاننده سلولهای اپی¬تلیال معدی رشد کرده و تشکیل کلنی میدهد، یافتههای آندوسکوپی خاصی نداشته و در نتیجه تشخیص بالینی آن مشکل است (15-13). با بررسیهای انجامشده، نقش هلیکوباکترپیلوری در ایجاد بدخیمی و سرطان معده تأیید شده است و همین امر موجب شده است که آژانس پژوهش سرطان سازمان بهداشت جهانی نام این باکتری را در ردیف عوامل سرطانزای کلاس I قرار دهد. تفاوت معناداری بین گسترش این عفونت در کشورهای غربی و کشورهای درحال توسعه به چشم میخورد. شیوع سرولوژی مثبت هلیکوباکترپیلوری در کشورهای درحال توسعه ۹۰ درصد جمعیت است، در حالیکه در کشورهای توسعه یافته، بهاستثنای ژاپن، شیوع زیر ۴۰ درصد است (16). این سرطان که مسئول مرگ ۶۵۰۰۰ نفر در جهان در سال ۲۰۰۰ بوده است، چهارمین بدخیمی رایج در دنیا است و ۱۰ درصد از کل مرگومیر سالیانه سرطان را به خود اختصاص میدهد (17). عفونت هلیکوباکترپیلوری در ایران نیز شایع بوده و این شیوع بین ۶۰ تا ۹۰ درصد است، بهخصوص سرطان معده از آمار بالایی برخوردار است (18). این مطلب نشان میدهد که ایران یک منطقه بسیار خطرناک برای عفونت هلیکوباکترپیلوری است. با توجه به اینکه هلیکوباکترپیلوری بهعنوان یک پاتوژن انسانی خطرناک به شمار میآید بنابراین ضرورت بررسی راههای پیشگیری از آن روشن است. در مسیر ابتلا به عفونت، آنتیژنهای این باکتری سبب شکلگیری واکنشهای دستگاه ایمنی میزبان میشوند بنابراین میتوان از ژنهای کدکننده این آنتیژنها بهصورت واکسنهای DNA بهره جست (19). واکسیناسیون هلیکوباکترپیلوری میتواند بار عفونت و پیامدهای آن را کاهش دهد. یک واکسن ایدهآل نهتنها باید ایمنی اثباتشده و سابقه بهرهوری خوبی داشته باشد بلکه باید ارزان قیمت بوده و ایمنی درازمدت ارائه دهد و کمترین تکرار دوز را نیاز داشته باشد. واکسیناسیون هلیکوباکترپیلوری، هم از منظر درمان این بیماری و هم از منظر جلوگیری از بروز آن، یک روش مناسب برای رفع این مشکل به نظر میآید (19). واکسنهای ژنی در مقایسه با سایر واکسنها دارای مزایایی چون تولید سادهتر، خالصتر بودن محصول، ذخیرهسازی و نگهداری آسان میباشند DNA. واکسن از یک پلاسمید تشکیلشده که درواقع یک DNA کوچک حلقوی است و میتواند در داخل سلول شروع به تکثیر کند، DNA واکسن به لحاظ ژنتیکی قابلیت دستکاری شدن را دارد و میتوان به کمک آن یک یا چندین آنتیژن اختصاصی از انواع مختلف عوامل عفونی و پاتوژنها را در سلول میزبان تولید و بیان کرد. وقتی DNA واکسن به سلول میزبان تزریق میشود میزبان شروع به خوانش از روی این پلاسمید کرده و پروتئینها و آنتیژنهای بیگانه در داخل سلول میزبان تولید میشود و این آنتیژنها توسط سلول میزبان پردازششده و در سطح سلول عرضه میگردد که بهاینترتیب سلولهای ایمنی از حضور آنها آگاه و سیستم ایمنی علیه آنها فعال میشود. همچنین وقتی سیستم ایمنی جهت پاسخ اولیه بر ضد آنتیژن بیگانه آماده میشود ایمنی محافظتی و خاطره¬ای نیز بر ضد آن پاتوژن تولید و منجر به مصونیت در مواجهه با پاتوژن میگردد (20). در پژوهش حاضر، قطعه ۱۷۵۷ جفت بازی ژن leoA با موفقیت از هلیکوباکترپیلوری استخراج و درون وکتور PTZ کلون و واکنش کلونینگ تأیید گردید. در مرحله بعد، قطعه درون وکتور بیانی pEGFP-C2 با موفقیت سابکلون گردید. پس از تأیید سابکلونینگ با آزمونهای تأییدی PCR و هضم آنزیمی، پروتئین نوترکیب ژن leoA در سلولهای CHO با وزن ۱۲۴ کیلودالتون بیان شد. نتایج حاصل از واکنش SDS-PAGE نشان داد که ژن leoA با موفقیت کلون شده است. سپس برای به دست آوردن مقادیر کافی از این پلاسمیدها، ابتدا با استفاده از باکتری E. coli سویه TOP10F، سلول مستعد تهیه شد و پلاسمیدهای مذکور بهصورت جداگانه به این سلولهای مستعد منتقل میشوند. ازآنجاکه پلاسمید pEGFP-C2-leoA، دارای ژن مقاومت به آنتیبیوتیک کانامایسین (بهعنوان مارکر انتخابی) است، در محیط کشت باکتریهای مستعد ترانسفورم شده با این پلاسمیدها، از آنتیبیوتیک کانامایسین (۱۰۰میلیگرم به ازای هر میلیلیتر محیط کشت باکتری) استفاده گردید. پس از رشد باکتریهای دارای پلاسمید، استخراج پلاسمید انجام گرفت و غلظت پلاسمیدهای تخلیص شده، با نانودراپ سنجیده شد. بهمنظور واکسیناسیون موشهای آزمایشگاهی نژاد BALB/c محلولهای تزریقی همانطور که توضیح داده شد ساخته و به عضله چهار سر ران در موشها تزریق شد. این تزریقات در طی ۳ مرحله (روزهای ۰، ۷ و ۱۵) انجام میشود. سپس سه گروه موش مختلف تعیین و به یک گروه پلاسمید نوترکیب به همراه نانو ذرات کیتوزان، به گروه دوم پلاسمید نوترکیب و به گروه سوم پلاسمید فاقد ژن کلون شده (گروه شاهد) تزریق گردید. پس از پانزده روز از آخرین مرحله تزریق واکسن، حیوانات آزمایشگاهی بهمنظور نمونهگیری تشریح شد، سپس خون آنها در تیوپ جمعآوری گردید. بافت مورد تزریق واکسن نیز جدا و در فریزر ۷۰- درجه سانتیگراد نگهداری شد. از خون (گلبولهای سفید) و بافت، استخراج RNA و سپس تهیه cDNA انجام گرفت. با استفاده از روش real time RT-PCR میزان بیان سایتوکاین¬هایی مثل IL-6، IL-4 و اینترفرون گاما بررسی شد و همچنین میزان بیان ژن leoA بهعنوان ژن بهکاررفته بهعنوان واکسن، در بافت مربوطه به همین روش بررسی گردید. از ژن GAPDH هم بهعنوان ژن رفرنس در کنار سایر آزمایشات استفاده شد. در نهایت اطلاعات بهدستآمده با استفاده از نرمافزار SPSS مورد بررسی آماری قرار گرفت. میاشیتا و همکاران در سال ۲۰۰۲ پاسخ ایمنی در موش بعد از واکسن ژنی کدکننده کاتالاز هلیکوباکترپیلوری را بررسی نمودند. در این آزمایش واکسن ژنی pcDNA3.1-kat استفاده شد، در این آزمایش موشهای C57/BL6 با ۱۰ میکروگرم ازpcDNA3.1-kat مورد آزمایش قرار گرفتند. آنتیبادی IGg مخصوص کاتالاز در سرم موشها مشاهده شد که باعث حفاظت آنها در برابر کلون شدن هلیکوباکترپیلوری گردید و سبب کاهش شدید درجه التهاب معده شد (21). مطالعهای در سال ۲۰۰۳ توسط اونیت همکارانش بهمنظور ارزیابی ایمنی در واکسن ژنی کدکننده سوپر اکسید دیسموتاز (SOD) انجام گرفت. با تزریق داخل عضلانی سازواره pcDNA-SOD به موشهای BALB/C انتقال یافت و سبب ایجاد پاسخ ایمنی هومورال و سلولی شد که سبب یک پاسخ ایمنی به Th-1 در موش و تولید اینترفرون گاما شد. این سازواره سطح قابلتوجهی از حفاظت در موش BALB/C در مقابل عفونت بروسلا آبورتوس نوع خطرناک ۲۳۰۵ القا کرده است. درمجموع این دادهها نشان داد که این سازواره کاندید خوبی برای استفاده در مطالعات آینده واکسیناسیون علیه بروسلوز است (22). در این مطالعه باکتری و نوع ژن موردبررسی با مطالعه ما متفاوت است. در سال ۲۰۰۴ در مطالعهای که توسط بایا و همکارانش انجام گرفت، ژن BabA2 توسط PCR تکثیر شد و وارد وکتور بیانی پروکاریوتی (+)pET-22b شد و در باکتری E. coli بیان شد. علاوه بر این، ایمنیزایی BabA با آزمایشهایی در حیوانات مورد بررسی قرار گرفت. پروتئین نوترکیب BabA سهمی حدود 34/8 درصد از کل پروتئین باکتریایی را به خود اختصاص داده بود. نتیجهگیری دست آمده از این مطالعه نشان داد که پروتئین نوترکیب BabA ممکن است یک واکسن بالقوه برای کنترل و درمان عفونت هلیکوباکترپیلوری باشد (23). در پژوهشی در سال ۲۰۰۵ کاساتارو و همکاران به ساخت واکسن ژنی علیه بروسلا پرداختند. در این پروژه پروتئین خارج سلولی ۳۱ (omp31) مورد توجه قرار گرفت و این پژوهشگران به بررسی اثرات این واکسن علیه B. melitensis و B. ovis پرداختند. Omp31 در وکتور بیانی pcI کلون شد و به موشهای BALB/C تزریق گردید. نتایج بهدستآمده نشان داد که این واکسن موجب واکنشهای سایتوتوکسیک میشود که توانایی مواجهه با عفونت بروسلا را دارد. این واکنشها به القای سلولهای CD8T میشود که سلولهای آلودهی بروسلا را از طریق مسیر پرفورین حذف میکند (24). این پژوهش ازلحاظ زیادی مشابه با پژوهش حاضر است تنها با این تفاوت که در مطالعه ما از ژن leoA استفاده شد. در پژوهش دیگری که در سال ۲۰۰۹ دوستی و همکاران با کلون سازی ژن omp31 در پی تولید واکسن ژنی برآمدند. نتایج این تحقیق و بررسی ایمنیزایی آن در موشهای BALB/C نشاندهنده فعالیت مناسب این واکسن ژنی بود که باعث واکنش سلولهای Th-1 در موشها گردید (25). در این مطالعه تنها ژن بررسیشده با مطالعه ما متفاوت است. در مطالعه دیگری در سال ۲۰۱۳ جای و همکاران اقدام به جداسازی و کلونسازی بخش ureI از مجموعه ژنهای اوره آزی نمودند. این محققان باهدف ساخت واکسن ژنی، ژن ureI را در وکتور بیانی PCDNA3.1 وارد نمودند و سازواره نهایی PCDNA3.1(+)-ureI را بهدست آوردند. این پژوهشگران نتیجه کار خود را اینگونه اعلام کردند که این سازواره قادر به بیان محصول پروتئینی ureI بوده و محرک سیستم ایمنی موشهای نژاد C57BL/6 بهصورت واکسن ژنی میباشد. نتیجهگیری این کار موفقیت این سازواره ژنی در ایجاد پاسخ ایمنی بسیار قوی از نوع سلولار و همولار بهطور همزمان بود (26). این مطالعه از چند جنبه مشابه و مؤید مطالعه حاضر است، هرچند نوع نژاد موشهای مورد استفاده متفاوت میباشد. در سال ۲۰۱۴ نیز تحقیق مشابه دیگری توسط المریری و همکاران انجام گرفت که به کلون سازی ژن B39 و Sp41 از بروسلا ملاتنسیز پرداختند و در پی تولید واکسن ژنی و بررسی ایمنیزایی آن در BALB/C به این نتیجه رسیدند که واکسن ژنی میتواند روش موفقی برای مبارزه علیه بروسلا باشد (27). این پژوهش نیز تا حدودی مشابه با مطالعه حاضر میباشد، هرچند نوع باکتری و ژن مورد نظر متفاوت میباشد. در پژوهشی که توسط محمودی واشیان و همکارش در سال ۲۰۱۶ انجام گرفت، نخست DNA هلیکوباکترپیلوری استخراج شد و ژن ureG با استفاده از پرایمر¬های ویژه تکثیر یافت. محصول PCR وارد وکتور PTZ شد و سپس با آنزیمهای SalI و xbaI برش داده شد و در وکتور بیانی pcI-neo ساب کلون گردید. سپس ureG -pcI-neo به روش الکتروپوریشن وارد سلول CHO کرده و بیان ژن ureG بر ژل SDS-PAGE مشاهده شد. نتایج نشان داد که ژن ureG کلونسازی شده توانایی بیان و تولید فرآورده پروتئینی اختصاصی این ژن در سلول جانوری CHO را دارد بنابراین این سازواره ژنی را میتوان بهعنوان کاندیدای مناسبی برای بررسیهای بعدی در مورد واکسنهای نوترکیب علیه هلیکوباکترپیلوری در الگوی حیوانی دانست. در این مطالعه ژنی از مجموعه اوره¬آز هلیکوباکتر مورد بررسی قرار گرفته است و از وکتور بیانی متفاوتی استفاده شده است (28).
نتیجهگیری
آنالیزهای آماری نشاندهنده افزایش سطح بیان سایتوکاین اینترفرون گاما و کاهش سطح بیان IL4 در منوسیتهای خون هر دو گروه از موشهای واکسینه شده بود. از طرفی سایتوکاین IL6 تغییر معنیداری در موشهای حاوی پلاسمید همراه ژن نشان نداد، درحالیکه در گروه حاوی پلاسمید همراه ژن همراه با نانو ذرات افزایش داشت. بهطورکلی این تغییرات نشاندهنده تحریک معنیدار سیستم ایمنی موشها میباشد. از طرف دیگر بررسی بیان ژن موردنظر در بافت عضله ران موشهای هر دو گروه در طی ۱۵، ۳۰ و ۴۵ روز پس از تزریق نشان داد که این بیان باگذشت زمان کاهش مییابد و تفاوت معنیداری هم در دو گروه موشهای واکسینه از این جنبه دیده نشد. روی هم رفته نتایج بهدستآمده نشاندهنده مؤثر بودن سازواره ژنی طراحیشده بهعنوان یک کاندید واکسن مناسب در درمان التهابات ناشی از هلیکوباکتر پیلوری ازجمله کارسینوم معده میباشد.
سپاسگزاری
نویسندگان این مقاله از معاونت علمی و فناوری دانشگاه آزاد اسلامی واحد فلاورجان و همکاران محترم مرکز تحقیقات بیوتکنولوژی دانشگاه آزاد اسلامی واحد شهرکرد به دلیل همکاری صمیمانه در اجرای این پژوهش کمال امتنان را دارند. و مطالعه حاضر منتج از پایاننامه است که با کد 1724812835810611400162428555 در دانشگاه آزاد اسلامی واحد فلاورجان تصویب شده است.
حامی مالی: ندارد.
تعارض در منافع: وجود ندارد.
References:
1- Suarez G, Reyes VE, Beswick EJ. Immune Response to H Pylori. World J Gastroenterology: WJG 2006; 12(35): 5593-8.
2- Meliț LE, Mărginean CO, Săsăran MO. The Challenges of Eradicating Pediatric Helicobacter Pylori Infection in the Era of Probiotics. Children 2022; 9(6): 795.
3- Lee A, O'Rourke J, De Ungria MC, Robertson B, Daskalopoulos G, Dixon MF. A standardized mouse model of Helicobacter pylori infection: introducing the Sydney strain. Gastroenterology 1997; 112(4): 1386-97.
4- Correa P. Human Gastric Carcinogenesis: A Multistep And Multifactorial Process—First American Cancer Society Award Lecture On Cancer Epidemiology And Prevention. Cancer Res 1992; 52(24): 6735-40.
5- Verbeke H, Geboes K, Van Damme J, Struyf S. The Role of CXC Chemokines in the Transition of Chronic Inflammation to Esophageal and Gastric Cancer. Biochim Biophys Acta 2013; 1835(2): 117-29.
6- Backert S, Naumann M. What a disorder: proinflammatory signaling pathways induced by Helicobacter pylori. Trends Microbiol 2010; 18(11): 479-86.
7- Koosirirat C, Linpisarn S, Changsom D, Chawansuntati K, Wipasa J. Investigation of the Anti-Inflammatory Effect of Curcuma Longa in Helicobacter Pylori-Infected Patients. International immunopharmacology 2010; 10(7): 815-8.
8- Rad R, Brenner L, Krug A, Voland P, Mages J, Lang R, et al. Toll-like receptor–dependent activation of antigen-presenting cells affects adaptive immunity to Helicobacter pylori. Gastroenterology 2007; 133(1): 150-63. e3.
9- Bengmark S, Mesa MD, Gil A. Plant-Derived Health-The Effects of Turmeric and Curcuminoids. Nutricion Hospitalaria 2009; 24(3): 273-81.
10- Yang YJ, Chen PC, Lai FP, Tsai PJ, Sheu BS. Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children. Biomedicines 2020; 8(6): 146.
11- Shakhatreh MAK, Khabour OF, Alzoubi KH, Banihani MN, Abu-Siniyeh A, Bashir NA, et al. The Influence of IL-1B Gene Polymorphisms on H. Pylori Infection and Triple Treatment Response among Jordanian Population. Appl Clin Genet 2020; 13: 139-45.
12- Hu Y, Zhang M, Lu B, Dai J. Helicobacter pylori and Antibiotic Resistance, A Continuing and Intractable Problem. Helicobacter 2016; 21(5): 349-63.
13- Nejati S, Karkhah A, Darvish H, Validi M, Ebarhimpour S, Nouri HR. Influence of Helicobacter Pylori Virulence Factors Caga and Vaca on Pathogenesis of Gastrointestinal Disorders. Microb Pathog 2018; 1; 117: 43-8.
14- Wu D, Cao M, Peng J, Li N, Yi S, Song L, et al. The Effect of Trimethylamine N-Oxide on Helicobacter Pylori-Induced Changes of Immunoinflammatory Genes Expression in Gastric Epithelial Cells. Int Immunopharmacol 2017; 43: 172-8.
15- Roszczenko-Jasińska P, Wojtyś MI, Jagusztyn-Krynicka EK. Helicobacter Pylori Treatment in the Post-Antibiotics Era—Searching for New Drug Targets. Appl Microbiol Biotechnol 2020; 104(23): 9891-905.
16- El-Shouny WA, Ali SS, Hegazy HM, Abd Elnabi MK, Ali A, Sun J. Syzygium Aromaticum L.: Traditional Herbal Medicine Against Caga and Vaca Toxin Genes-Producing Drug Resistant Helicobacter Pylori. J Tradit Complement Med 2019; 10(4): 366-77.
17- Banga Ndzouboukou JL, Lei Q, Ullah N, Zhang Y, Hao L, Fan X. Helicobacter pylori adhesins: HpaA a potential antigen in experimental vaccines for H. pylori. Helicobacter 2021; 26(1): e12758.
18- Mirzaei N, Poursina F, Moghim S, Rashidi N, Ghasemian Safaei H. The Study of H. Pylori Putative Candidate Factors for Single- and Multi-Component Vaccine Development. Crit Rev Microbiol 2017; 43(5): 631-50.
19- Aliramaei MR, Khorasgani MR, Rahmani MR, Zarkesh Esfahani SH, Emamzadeh R. Expression of Helicobacter Pylori Cagl Gene In Lactococcus Lactis MG1363 and Evaluation of Its Immunogenicity as an Oral Vaccine in Mice. Microb Pathog 2020; 142: 103926.
20- Pan X, Ke H, Niu X, Li S, Lv J, Pan L. Protection Against Helicobacter Pylori Infection in BALB/C Mouse Model ذy Oral Administration of Multivalent Epitope-Based Vaccine of Cholera Toxin B Subunit-HUUC. Front Immunol 2018; 9: 1003.
21- Czinn SJ, Blanchard T. Vaccinating Against Helicobacter Pylori Infection. Nat Rev Gastroenterol Hepatol 2011; 8(3): 133-40.
22- Miyashita M, Joh T, Watanabe K, Todoroki I, Seno K, Ohara H, et al. Immune Responses in Mice to Intranasal and Intracutaneous Administration of a DNA Vaccine Encoding Helicobacter Pylori-Catalase. Vaccine 2002; 20(17-18): 2336-42.
23- Oñate AA, Céspedes S, Cabrera A, Rivers R, González A, Muñoz C, et al. A DNA Vaccine Encoding Cu,Zn Superoxide Dismutase Of Brucella Abortus Induces Protective Immunity In BALB/C Mice. Infect Immun 2003; 71(9): 4857-61.
24- Bai Y, Zhang YL, Chen Y, Jin JF, Zhang ZS, Zhou DY. Cloning and Expression and Immunogenicity of Helicobacter Pylori Baba2 Gene. World J Gastroenterol 2004; 10(17): 2560-2.
25- Cassataro J, Velikovsky CA, De La Barrera S, Estein SM, Bruno L, Bowden R, et al. A DNA vaccine coding for the Brucella outer membrane protein 31 confers protection against B. melitensis and B. Ovis Infection by Eliciting a Specific Cytotoxic Response. Infect Immun 2005; 73(10): 6537-46.
26- Doosti A, Ghasemi-Dehkordi P, Javadi GR, Sardari S, Shokrgozar MA. DNA Vaccine Encoding The Omp31 Gene Of Brucella Melitensis Induces Protective Immunity In BALB/C Mice. Res J Biol Sci 2009; 4(1): 126-31.
27- Zhao YX, Qi JH, Zhang H, Duan GC, Xi YL. Construction, Expression and Purification of Ureb-Omp11 Fusion Protein of Helicobacter Pylori and its Immunocompetence. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2007; 23(10): 906-10.
28- Al-Mariri A, Akel R, Abbady AQ. A DNA Vaccine Encoding P39 and Sp41 of Brucella Melitensis Induces Protective Immunity in BALB/C Mice. Arch Med Vet 2014; 46(1): 53-62.
هلیکوباکتر پیلوری (H. pylori) یکی از شایعترین پاتوژنهای انسانی است زیرا مخاط معده حدود 50 درصد از جمعیت جهان را آلوده میکند (1). اکثر عفونتهای ایجاد شده بدون علامت هستند و بیشتر در جمعیت اطفال ایجاد میشوند که عفونت را بدون ریشهکنی مؤثر باکتریایی مادامالعمر میسازد. علاوه بر این، مطالعات اپیدمیولوژیک که عفونت را با خطر بالاتر بدخیمی معده مرتبط میکند، سازمان بهداشت جهانی را برای تحقیقات سرطان جهت طبقهبندی هلیکوباکتر پیلوری بهعنوان سرطانزای کلاس I هدایت میکند (2). عفونت هلیکوباکتر پیلوری یک عامل خطر اصلی برای توسعه سرطان معده است زیرا باعث ایجاد یک توالی گامبهگام در مخاط معده میشود که با گاستریت سطحی شروع میشود، که میتواند به گاستریت مزمن، گاستریت آتروفیک، متاپلازی روده، دیسپلازی و درنهایت کارسینوم معده تبدیل شود (4،3). باکتریها پاسخ ایمنی میزبان (ذاتی و سازگار) را القا میکنند، اما تداوم عفونت نشان میدهد که پاسخ در از بین بردن عفونت مؤثر نیست. علاوه بر این، شواهد متعدد نشان میدهد که پاسخ ایمنی به پاتوژنز مرتبط با عفونت کمک میکند. در طول دو دهه گذشته، چندین مدل تجربی از عفونت هلیکوباکتر پیلوری برای بررسی پاتوژنز این عفونت ایجادشده است. با استفاده از سویه هلیکوباکتر پیلوری سازگار با موش (Sidney Strain،SS1)، لی و همکاران. مدلی از کلونیزاسیون باکتریایی طولانیمدت و بالا را در موشها ایجاد کردند (3). اگرچه هلیکوباکتر پیلوری بهعنوان سویه غیرتهاجمی شناخته شده است، اما باعث ایجاد یک واکنش التهابی گسترده در مخاط معده میشود (5). این واکنش با نفوذ مخاطی سلولهای التهابی، بهویژه نوتروفیلها، مشخص میشود که با افزایش بیان کموکاینها و سیتوکینهای پیش التهابی انجام میشود (7،6). با توجه به شیوع بالای عفونت هلیکوباکتر پیلوری در سراسر جهان، هزینههای بالای درمان آنتیبیوتیکی و افزایش نرخ مقاومت آنتیبیوتیکی، تلاشهای قابلتوجهی برای تولید واکسن علیه هلیکوباکتر پیلوری انجام شده است. بااینحال، واکسیناسیون کارآمد تا به امروز در انسان به دست نیامده است (8). بنابراین، یافتن درمانهای جایگزین ضروری است (9). ژن leoA یکی از عوامل بیماریزایی GTPase هلیکوباکترپیلوری است که در جزیره بیماریزایی کدگذاری میشود و بالقوه از طریق وزیکول ترشحی باعـث افـزایش انتشـار سـم مـیشـود (10). وزیکولهای غشاء نقش مهمـی در عملکـرد حفاظتشده اعضـای خانواده دیآمینی، دارند. گزارشات نشان دادهاند که این ژن توانایی ایمنیزایی بالقوه در بدن میزبان را دارد؛ لذا آن را نامزد مناسبی برای تولید یک واکسن ژنی علیه هلیکوباکترپیلوری، معرفی نمودهاند. هدف از این مطالعه بررسی میزان بیان ژنهای IL6، IL4 و اینترفرون گاما در سطح mRNA به روش real time PCR در خون موشهای BALB/c واکسینه شده با واکسن ژنی pEGFP-C2-leoA بوده است. همچنین میزان بیان و ماندگاری leoA در بافت محل تزریق واکسن (عضله ران موشها) در یک دوره زمانی ۴۵ روزه بررسیشده است.
روش بررسی
کلونسازی ژن در وکتور بیانی: در این مطالعه، قطعه ژن leoA هلیکوباکترپیلوری درون وکتور بیانی pEGFP-C2 طراحی و توسط شرکت Generay سنتز گردید. باکتریE. coli سویه Top10F بهمنظور ترانسفورماسیون و تکثیر سازههای ژنی نوترکیب مورداستفاده قرار گرفت. انتخاب و غربالگری باکتریهای دریافتکننده پلاسمید نوترکیب در حضور آنتیبیوتیک کانامایسین انجام شد. صحت وکتور نوترکیب (pEGFP-C2-leoA) به روش PCR و هضم آنزیمی با آنزیمهای KpnI و SacII تأیید گردید. همچنین صحت توالی ژن کلون شده درون وکتور بیانی pEGFP-C2، توسط شرکت سازنده و با روش تعیین توالی سانگر موردبررسی قرار گرفت.
انتقال سازواره نهایی pEGFP-C2-leoA به سلولهای جانوری: بهمنظور بررسی بیان ژن leoA در سلول جانوری، از سلول CHO استفاده شد و برای ترانسفورمیشن این سلولها از روش الکتروپوریشن (مدل Gene Pulser Xcell ساخت شرکت Bio Rad) بهره گرفته شد. تعداد دو میلیون سلولهای CHO به همراه ۸۰۰ نانوگرم در هر میکرو لیتر از وکتور نوترکیب pEGFP-C2-leoA و وکتور pEGFP-C2 بدون ژن در دو گروه مختلف در حجم ۴۰۰ میکرولیتر در کوت 0/4 میلیمتری ریخته شد. پالس الکتریکی با شرایط بهینهسازی شده 0/174 کیلوولت و ۴۰۰ میکرو فاراد به سلولها داده شد و سلولهای حاصل در حضور آنتیبیوتیک نئومایسین کشت داده شدند.
انجام SDS-PAGE: وزن مولکولی و محل قرارگرفته پروتئین بیانشده درون سلولهای یوکاریوتی CHO، بهوسیله آزمون SDS-PAGE تأیید شد. سلولهای CHO ترانسفورم شده به مدت ۳ دقیقه با دور ۳۰۰۰ دور بر دقیقه رسوب داده شدند. به رسوب سلولی مقدار ۱۰۰ میکرو لیتر PBS اضافه شد و در نهایت سلولهای حاصل بر روی ژل ۱۲ درصد SDS-PAGE الکتروفورز و با کوماسی بلو رنگآمیزی انجام گرفت.
تهیه نانو ذرات کیتوزان به روش Ionic Gelation: ابتدا محلول کیتوزان (حلکردن صد میلیگرم کیتوزان در ۵۰ میلیلیتر اسید استیک به مدت ۲۴ ساعت در دمای اتاق با ۵/۵= pH و فیلتر 45/0 میکرومتری جهت حذف کیتوزانهای حلنشده) تهیه شد. سپس محلول TPP (۲۰میلیگرم در ۲۰ میلیلیتر آب دیونیزه) بهصورت قطرهقطره به محلول کیتوزان در حال چرخش روی همزن در مدت ۱ ساعت با دور rpm ۱۰۰۰ اضافه شد. سپس محلول حاصل به مدت ۱۵ دقیقه با دور rpm ۱۴۰۰۰سانتریفیوژ شد و محلول رویی حاوی نانو ذرات کیتوزان جمعآوری گردید و با دستگاه فریز - درایر خشک و پودر گردید. از روشهای زتا آنالایزر (دستگاه Nanozeta Sizer ساخت کشور آمریکا) برای ارزیابی ویژگی-های فیزیکوشیمیایی، شاخص پراکندگی و اندازه نانو ذرات کیتوزان تولیدشده استفاده شد و از دستگاه Malvern Instruments ساخت کشور انگلستان و میکروسکوپ الکترونی (SEM) برای پراکندگی نوری دینامیک (Dynamic Light Scattering) استفاده گردید. تهیه مخلوط پلاسمید و نانو ذرات: نسبت مساوی از محلول کیتوزان (۱ درصد) و پلاسمید (۲۰۰۰ میکروگرم در هر میلیلیتر PBS) باهم مخلوط شدند و به مدت ۱ ساعت در دمای ۵۵ درجه سانتیگراد قرار گرفتند.
گروهبندی و زمانبندی تزریق به موشها
18 سر موشهای ماده BALB/c ششهفتهای با وزن 30-22 گرم به سه گروه ۶تایی دستهبندی شدند:
- گروه اول: پلاسمید نوترکیب (واکسن ژنی) + نانو ذرات کیتوزان
- گروه دوم: پلاسمید نوترکیب (واکسن ژنی)
- گروه سوم: پلاسمید فاقد ژن هدف (گروه شاهد)
تزریق گردید. تزریقها بافاصله زمانی ۰، ۷ و ۱۵ روز انجام گرفت.
تزریق به موش: موشهای گروه اول مقدار ۱۰۰ میکرولیتر (حاوی ۱۰۰ میکروگرم DNA)، و موشهای گروه دوم ۱۰۰ میکرولیتر (حاوی ۱۰۰۰ میکروگرم در هر میلیلیتر PBS را بهصورت تزریقی دریافت کردند. به گروه شاهد نیز محلولی بهصورت ۱۰۰۰ میکروگرم در هر میلیلیتر PBS تهیه شد. سپس ۱۰۰ میکرولیتر از آن تزریق گردید.
نمونهبرداری: در روزهای پانزدهم، سیام و چهل و پنجم پس از آخرین مرحله تزریق، تعداد ۲ سر موش از هر گروه، کشته و نمونهبرداری شد.
بافت عضله ران موش) دقیقاً در محل تزریق(، و مونوسیتهای خون کامل با استفاده از فایکول جداسازی شد.
استخراج RNA: با استفاده از کیت استخراج RNA (شرکت یکتا تجهیز)، RNA کلی بافت طبق دستورالعمل شرکت سازنده تخلیص شد و آن با استفاده از الکتروفورز روی ژل آگارز ۱ درصد و مشاهده باندهای 18S و 28S بررسی گردید. همچنین غلظت RNAها با استفاده از دستگاه نانودراپ سنجیده شد. بلافاصله جهت سنتز cDNA مورداستفاده قرار گرفت.
ساخت Cdna: برای سنتزcDNA ، یک میکروگـرم از RNA استخراجشده با اسـتفاده از آغـازگر الیگـومر تیمیدین (Oligo dT) و کیـت شــرکت تاکارا ژاپن، در حجم ۱۰ میکرولیتر طبـق دستورالعمل شــرکت ســازنده بــه cDNA تبدیل گردید.
واکنش RT-qPCR: پرایمرهای مورداستفاده توسط نرمافزار Oligo 7 و برنامه Blast طراحی شدند (جدول ۱). هر واکـنش PCR (شامل SYBR Green Master Mix، پرایمر، cDNA و آب مقطر) در ۴۵ سیکل انجام شد و از ژن GAPDH بهعنوان ژن رفرنس در تمام آزمایشات بیان ژن استفاده شد.
تجزیهوتحلیل آماری
نتایج بهدستآمده بهوسیله نرمافزار SPSS version 16 مورد تحلیل آماری قرارگرفت؛ از آزمون one way ANOVAو آزمون متعاقب LSD و نیز آزمونمستقل t-test ، جهت بررسی و وجود ارتباط و میزان معنیداری دادهها استفاده گردید. تمام دادهها بهصورت means + S.E.M در سطح معنیداری p<0.05 در نظر گرفته شدند. آنالیز دادهها با استفاده از متد CtΔΔ-۲ انجام شد. مزیت روش ΔΔCT در سهولت مراحل کار است و یکبار رسم منحنی استاندارد کافی میباشد و در صورتی از این روش استفاده میشود که کارایی PCR در نمونههای کنترل و هدف نزدیک به 100٪ باشد.
ملاحظات اخلاقی
مطالعه حاضر از نوع تجربی میباشد که در کمیته اخلاق دانشگاه آزاد اسلامی واحد فلاورجان با کدIR.IAU.FALA.REC.1400.023 به تصویب رسیده است.
جدول 1: توالی پرایمر استفادهشده در این تحقیق
نتایج
صحت کلون شدن ژن leoA: درستی سازواره طراحیشده با روشهای PCR و هضم آنزیمی (KpnI و SacII) مورد تأیید قرار گرفت. برش وکتور pEGFP-C2-leoA با دو آنزیم فوقالذکر سبب تشکیل دو قطعه، با اندازههای ۶۴۹۲ جفت باز و ۱۷۵۷ جفت باز گردید که نشاندهنده تشکیل سازواره نهایی بود (شکل ۱).
الکتروپوریشن و بیان ژن: درستی ورود سازواره pEGFP-C2-leoA در سلولهای CHO با واکنش PCR و رشد در حضور آنتیبیوتیک تأیید شد. بیان ژن leoA در این سلولها، با واکنش SDS-PAGE انجام شد. محصول پروتئینی حاصل از ژن leoA به وزن ۱۲۴ کیلو دالتون نشاندهنده بیان این پروتئین در سلولهای یوکاریوتی CHO بود (شکل ۱).
تأیید نانو ذرات کیتوزان: اندازه نانو ذرات کیتوزان (وزن مولکولی پایین) با استفاده از میکروسکوپ الکترونی کمتر از ۲۰۰ نانومتر به دست آمد و میانگین اندازه نانو ذرات بهدست آمده توسط پراکنگی نوری دینامیک (DLS) 111/7 نانومتر و همچنین شاخص پراکنگی اندازه ذرات توسط آنالیز زتا (mV20/8) تعیین شد.
بررسی بیان ژنها: ژن leoA: بیان این ژن در نمونه بافت بهدستآمده از عضله ران موشهای واکسینه شده به روشreal time RT-PCR انجام شد. منحنیهای ذوب مربوط به ژنهای leoA و GAPDH بهصورت تک قله به دست آمد که بیانگر وجود تنهـا یـک محـصول خاص در PCR است. مقایسه بیان ژن leoA در بافت موشهای گروه حاوی پلاسمید با گروه حاوی نانو ذرات کیتوزان در روز ۱۵ افزایش بیان داشت و در روز ۴۵ کاهش بیان را نشان داد (نمودار ۱).
بیان ژنهای IL6، γIFN و IL4: بیان این ژنها در نمونه خون موشهای واکسینه شده با روش real time RT-PCR اندازهگیری شد. منحنیهای ذوب مربوط به ژنهای IL6، IFNᵧ، و IL4بهصورت تک قله به دست آمد که بیانگر وجود یـک محـصول خاص درPCR است. مقایسه میانگین بیان نسبی این ژنها نشان داد IFNᵧ در نمونههای پلاسمید و نمونه پلاسمید به همراه نانو ذرات کیتوزان نسبت به نرمال، افزایش بیان دارد. همچنین IL6 در نمونههای پلاسمید و نمونههای پلاسمید به همراه نانو ذرات کیتوزان نسبت به نرمال، کاهش بیان نسبت به نرمال نشان داد (نمودار ۱).
شکل 1: A) تکثیر ژن leoA به روش PCR، B) هضم آنزیمی سازواره نهایی pEGFP-C2-leoA با دو آنزیم KpnI و SacII و C) محصول پروتئینی حاصل از ژن leoA
نمودار 1: A: بررسی بیان ژن leoA در بافت موشهای حاوی پلاسمید همراه با ژن که در زمانهای 15، 30 و 45 بهصورت معنیدار (P <.0.01) اختلاف دارند. B) بررسی بیان ژن leoA در بافت موشهای حاوی پلاسمید همراه با ژن همراه با نانوذره کیتوزان که در زمانهای 15، 30 و 45 بهصورت معنیدار (P<.0.01) اختلاف دارند. C) بررسی بیان سایتوکاینها در مونوسیت استخراجشده از خون کامل موشهای حاوی پلاسمید همراه با ژن همراه با نانوذره که در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IFNᵧ بهصورت معنیدار افزایشیافته است (P<.0.01). در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IL6 بهصورت معنیدار افزایشیافته است (P<.0.5). در گروه حاوی pEGFP-C2-leoA + Nanoparticle بیان ژن IL4 بهصورت معنیدار کاهشیافته است (P<.0.01). D) بررسی بیان سایتوکاینها مونوسیت استخراجشده از خون کامل موشهای حاوی پلاسمید همراه با ژن که در گروه pEGFP-C2-leoA بیان ژن IFNᵧ بهصورت معنیدار (P<.001) افزایشیافته است، در گروه pEGFP-C2-leoA بیان ژن IL6 بهصورت معنیدار (P<0.5) تغییر نداشته است و در گروه pEGFP-C2-leoA بیان ژن IL4 بهصورت معنیدار (P<.0.01) کاهشیافته است.
بحث
هلیکوباکترپیلوری یک باکتری گرم منفی، متحرک، میلهای شکل و میکروآئروفیل (باکتریهایی که اکسیژن را بهعنوان گیرنده نهایی الکترون نیاز دارند و در شرایط بیهوازی به مقدار خیلی کمی رشد میکنند) است، که در سطح مجرایی پوشش معده دیده میشود (11). این باکتری دلیل اصلی گاستریت فعال و مزمن، زخم معده و دوازدهه است. همچنین موجب التهاب طولانی در مخاط معده بهوسیله نوتروفیلها، لنفوسیتها و پلاسماسلها میشود (12). ازآنجاییکه این باکتری در لایه عمقی موکوس پوشاننده سلولهای اپی¬تلیال معدی رشد کرده و تشکیل کلنی میدهد، یافتههای آندوسکوپی خاصی نداشته و در نتیجه تشخیص بالینی آن مشکل است (15-13). با بررسیهای انجامشده، نقش هلیکوباکترپیلوری در ایجاد بدخیمی و سرطان معده تأیید شده است و همین امر موجب شده است که آژانس پژوهش سرطان سازمان بهداشت جهانی نام این باکتری را در ردیف عوامل سرطانزای کلاس I قرار دهد. تفاوت معناداری بین گسترش این عفونت در کشورهای غربی و کشورهای درحال توسعه به چشم میخورد. شیوع سرولوژی مثبت هلیکوباکترپیلوری در کشورهای درحال توسعه ۹۰ درصد جمعیت است، در حالیکه در کشورهای توسعه یافته، بهاستثنای ژاپن، شیوع زیر ۴۰ درصد است (16). این سرطان که مسئول مرگ ۶۵۰۰۰ نفر در جهان در سال ۲۰۰۰ بوده است، چهارمین بدخیمی رایج در دنیا است و ۱۰ درصد از کل مرگومیر سالیانه سرطان را به خود اختصاص میدهد (17). عفونت هلیکوباکترپیلوری در ایران نیز شایع بوده و این شیوع بین ۶۰ تا ۹۰ درصد است، بهخصوص سرطان معده از آمار بالایی برخوردار است (18). این مطلب نشان میدهد که ایران یک منطقه بسیار خطرناک برای عفونت هلیکوباکترپیلوری است. با توجه به اینکه هلیکوباکترپیلوری بهعنوان یک پاتوژن انسانی خطرناک به شمار میآید بنابراین ضرورت بررسی راههای پیشگیری از آن روشن است. در مسیر ابتلا به عفونت، آنتیژنهای این باکتری سبب شکلگیری واکنشهای دستگاه ایمنی میزبان میشوند بنابراین میتوان از ژنهای کدکننده این آنتیژنها بهصورت واکسنهای DNA بهره جست (19). واکسیناسیون هلیکوباکترپیلوری میتواند بار عفونت و پیامدهای آن را کاهش دهد. یک واکسن ایدهآل نهتنها باید ایمنی اثباتشده و سابقه بهرهوری خوبی داشته باشد بلکه باید ارزان قیمت بوده و ایمنی درازمدت ارائه دهد و کمترین تکرار دوز را نیاز داشته باشد. واکسیناسیون هلیکوباکترپیلوری، هم از منظر درمان این بیماری و هم از منظر جلوگیری از بروز آن، یک روش مناسب برای رفع این مشکل به نظر میآید (19). واکسنهای ژنی در مقایسه با سایر واکسنها دارای مزایایی چون تولید سادهتر، خالصتر بودن محصول، ذخیرهسازی و نگهداری آسان میباشند DNA. واکسن از یک پلاسمید تشکیلشده که درواقع یک DNA کوچک حلقوی است و میتواند در داخل سلول شروع به تکثیر کند، DNA واکسن به لحاظ ژنتیکی قابلیت دستکاری شدن را دارد و میتوان به کمک آن یک یا چندین آنتیژن اختصاصی از انواع مختلف عوامل عفونی و پاتوژنها را در سلول میزبان تولید و بیان کرد. وقتی DNA واکسن به سلول میزبان تزریق میشود میزبان شروع به خوانش از روی این پلاسمید کرده و پروتئینها و آنتیژنهای بیگانه در داخل سلول میزبان تولید میشود و این آنتیژنها توسط سلول میزبان پردازششده و در سطح سلول عرضه میگردد که بهاینترتیب سلولهای ایمنی از حضور آنها آگاه و سیستم ایمنی علیه آنها فعال میشود. همچنین وقتی سیستم ایمنی جهت پاسخ اولیه بر ضد آنتیژن بیگانه آماده میشود ایمنی محافظتی و خاطره¬ای نیز بر ضد آن پاتوژن تولید و منجر به مصونیت در مواجهه با پاتوژن میگردد (20). در پژوهش حاضر، قطعه ۱۷۵۷ جفت بازی ژن leoA با موفقیت از هلیکوباکترپیلوری استخراج و درون وکتور PTZ کلون و واکنش کلونینگ تأیید گردید. در مرحله بعد، قطعه درون وکتور بیانی pEGFP-C2 با موفقیت سابکلون گردید. پس از تأیید سابکلونینگ با آزمونهای تأییدی PCR و هضم آنزیمی، پروتئین نوترکیب ژن leoA در سلولهای CHO با وزن ۱۲۴ کیلودالتون بیان شد. نتایج حاصل از واکنش SDS-PAGE نشان داد که ژن leoA با موفقیت کلون شده است. سپس برای به دست آوردن مقادیر کافی از این پلاسمیدها، ابتدا با استفاده از باکتری E. coli سویه TOP10F، سلول مستعد تهیه شد و پلاسمیدهای مذکور بهصورت جداگانه به این سلولهای مستعد منتقل میشوند. ازآنجاکه پلاسمید pEGFP-C2-leoA، دارای ژن مقاومت به آنتیبیوتیک کانامایسین (بهعنوان مارکر انتخابی) است، در محیط کشت باکتریهای مستعد ترانسفورم شده با این پلاسمیدها، از آنتیبیوتیک کانامایسین (۱۰۰میلیگرم به ازای هر میلیلیتر محیط کشت باکتری) استفاده گردید. پس از رشد باکتریهای دارای پلاسمید، استخراج پلاسمید انجام گرفت و غلظت پلاسمیدهای تخلیص شده، با نانودراپ سنجیده شد. بهمنظور واکسیناسیون موشهای آزمایشگاهی نژاد BALB/c محلولهای تزریقی همانطور که توضیح داده شد ساخته و به عضله چهار سر ران در موشها تزریق شد. این تزریقات در طی ۳ مرحله (روزهای ۰، ۷ و ۱۵) انجام میشود. سپس سه گروه موش مختلف تعیین و به یک گروه پلاسمید نوترکیب به همراه نانو ذرات کیتوزان، به گروه دوم پلاسمید نوترکیب و به گروه سوم پلاسمید فاقد ژن کلون شده (گروه شاهد) تزریق گردید. پس از پانزده روز از آخرین مرحله تزریق واکسن، حیوانات آزمایشگاهی بهمنظور نمونهگیری تشریح شد، سپس خون آنها در تیوپ جمعآوری گردید. بافت مورد تزریق واکسن نیز جدا و در فریزر ۷۰- درجه سانتیگراد نگهداری شد. از خون (گلبولهای سفید) و بافت، استخراج RNA و سپس تهیه cDNA انجام گرفت. با استفاده از روش real time RT-PCR میزان بیان سایتوکاین¬هایی مثل IL-6، IL-4 و اینترفرون گاما بررسی شد و همچنین میزان بیان ژن leoA بهعنوان ژن بهکاررفته بهعنوان واکسن، در بافت مربوطه به همین روش بررسی گردید. از ژن GAPDH هم بهعنوان ژن رفرنس در کنار سایر آزمایشات استفاده شد. در نهایت اطلاعات بهدستآمده با استفاده از نرمافزار SPSS مورد بررسی آماری قرار گرفت. میاشیتا و همکاران در سال ۲۰۰۲ پاسخ ایمنی در موش بعد از واکسن ژنی کدکننده کاتالاز هلیکوباکترپیلوری را بررسی نمودند. در این آزمایش واکسن ژنی pcDNA3.1-kat استفاده شد، در این آزمایش موشهای C57/BL6 با ۱۰ میکروگرم ازpcDNA3.1-kat مورد آزمایش قرار گرفتند. آنتیبادی IGg مخصوص کاتالاز در سرم موشها مشاهده شد که باعث حفاظت آنها در برابر کلون شدن هلیکوباکترپیلوری گردید و سبب کاهش شدید درجه التهاب معده شد (21). مطالعهای در سال ۲۰۰۳ توسط اونیت همکارانش بهمنظور ارزیابی ایمنی در واکسن ژنی کدکننده سوپر اکسید دیسموتاز (SOD) انجام گرفت. با تزریق داخل عضلانی سازواره pcDNA-SOD به موشهای BALB/C انتقال یافت و سبب ایجاد پاسخ ایمنی هومورال و سلولی شد که سبب یک پاسخ ایمنی به Th-1 در موش و تولید اینترفرون گاما شد. این سازواره سطح قابلتوجهی از حفاظت در موش BALB/C در مقابل عفونت بروسلا آبورتوس نوع خطرناک ۲۳۰۵ القا کرده است. درمجموع این دادهها نشان داد که این سازواره کاندید خوبی برای استفاده در مطالعات آینده واکسیناسیون علیه بروسلوز است (22). در این مطالعه باکتری و نوع ژن موردبررسی با مطالعه ما متفاوت است. در سال ۲۰۰۴ در مطالعهای که توسط بایا و همکارانش انجام گرفت، ژن BabA2 توسط PCR تکثیر شد و وارد وکتور بیانی پروکاریوتی (+)pET-22b شد و در باکتری E. coli بیان شد. علاوه بر این، ایمنیزایی BabA با آزمایشهایی در حیوانات مورد بررسی قرار گرفت. پروتئین نوترکیب BabA سهمی حدود 34/8 درصد از کل پروتئین باکتریایی را به خود اختصاص داده بود. نتیجهگیری دست آمده از این مطالعه نشان داد که پروتئین نوترکیب BabA ممکن است یک واکسن بالقوه برای کنترل و درمان عفونت هلیکوباکترپیلوری باشد (23). در پژوهشی در سال ۲۰۰۵ کاساتارو و همکاران به ساخت واکسن ژنی علیه بروسلا پرداختند. در این پروژه پروتئین خارج سلولی ۳۱ (omp31) مورد توجه قرار گرفت و این پژوهشگران به بررسی اثرات این واکسن علیه B. melitensis و B. ovis پرداختند. Omp31 در وکتور بیانی pcI کلون شد و به موشهای BALB/C تزریق گردید. نتایج بهدستآمده نشان داد که این واکسن موجب واکنشهای سایتوتوکسیک میشود که توانایی مواجهه با عفونت بروسلا را دارد. این واکنشها به القای سلولهای CD8T میشود که سلولهای آلودهی بروسلا را از طریق مسیر پرفورین حذف میکند (24). این پژوهش ازلحاظ زیادی مشابه با پژوهش حاضر است تنها با این تفاوت که در مطالعه ما از ژن leoA استفاده شد. در پژوهش دیگری که در سال ۲۰۰۹ دوستی و همکاران با کلون سازی ژن omp31 در پی تولید واکسن ژنی برآمدند. نتایج این تحقیق و بررسی ایمنیزایی آن در موشهای BALB/C نشاندهنده فعالیت مناسب این واکسن ژنی بود که باعث واکنش سلولهای Th-1 در موشها گردید (25). در این مطالعه تنها ژن بررسیشده با مطالعه ما متفاوت است. در مطالعه دیگری در سال ۲۰۱۳ جای و همکاران اقدام به جداسازی و کلونسازی بخش ureI از مجموعه ژنهای اوره آزی نمودند. این محققان باهدف ساخت واکسن ژنی، ژن ureI را در وکتور بیانی PCDNA3.1 وارد نمودند و سازواره نهایی PCDNA3.1(+)-ureI را بهدست آوردند. این پژوهشگران نتیجه کار خود را اینگونه اعلام کردند که این سازواره قادر به بیان محصول پروتئینی ureI بوده و محرک سیستم ایمنی موشهای نژاد C57BL/6 بهصورت واکسن ژنی میباشد. نتیجهگیری این کار موفقیت این سازواره ژنی در ایجاد پاسخ ایمنی بسیار قوی از نوع سلولار و همولار بهطور همزمان بود (26). این مطالعه از چند جنبه مشابه و مؤید مطالعه حاضر است، هرچند نوع نژاد موشهای مورد استفاده متفاوت میباشد. در سال ۲۰۱۴ نیز تحقیق مشابه دیگری توسط المریری و همکاران انجام گرفت که به کلون سازی ژن B39 و Sp41 از بروسلا ملاتنسیز پرداختند و در پی تولید واکسن ژنی و بررسی ایمنیزایی آن در BALB/C به این نتیجه رسیدند که واکسن ژنی میتواند روش موفقی برای مبارزه علیه بروسلا باشد (27). این پژوهش نیز تا حدودی مشابه با مطالعه حاضر میباشد، هرچند نوع باکتری و ژن مورد نظر متفاوت میباشد. در پژوهشی که توسط محمودی واشیان و همکارش در سال ۲۰۱۶ انجام گرفت، نخست DNA هلیکوباکترپیلوری استخراج شد و ژن ureG با استفاده از پرایمر¬های ویژه تکثیر یافت. محصول PCR وارد وکتور PTZ شد و سپس با آنزیمهای SalI و xbaI برش داده شد و در وکتور بیانی pcI-neo ساب کلون گردید. سپس ureG -pcI-neo به روش الکتروپوریشن وارد سلول CHO کرده و بیان ژن ureG بر ژل SDS-PAGE مشاهده شد. نتایج نشان داد که ژن ureG کلونسازی شده توانایی بیان و تولید فرآورده پروتئینی اختصاصی این ژن در سلول جانوری CHO را دارد بنابراین این سازواره ژنی را میتوان بهعنوان کاندیدای مناسبی برای بررسیهای بعدی در مورد واکسنهای نوترکیب علیه هلیکوباکترپیلوری در الگوی حیوانی دانست. در این مطالعه ژنی از مجموعه اوره¬آز هلیکوباکتر مورد بررسی قرار گرفته است و از وکتور بیانی متفاوتی استفاده شده است (28).
نتیجهگیری
آنالیزهای آماری نشاندهنده افزایش سطح بیان سایتوکاین اینترفرون گاما و کاهش سطح بیان IL4 در منوسیتهای خون هر دو گروه از موشهای واکسینه شده بود. از طرفی سایتوکاین IL6 تغییر معنیداری در موشهای حاوی پلاسمید همراه ژن نشان نداد، درحالیکه در گروه حاوی پلاسمید همراه ژن همراه با نانو ذرات افزایش داشت. بهطورکلی این تغییرات نشاندهنده تحریک معنیدار سیستم ایمنی موشها میباشد. از طرف دیگر بررسی بیان ژن موردنظر در بافت عضله ران موشهای هر دو گروه در طی ۱۵، ۳۰ و ۴۵ روز پس از تزریق نشان داد که این بیان باگذشت زمان کاهش مییابد و تفاوت معنیداری هم در دو گروه موشهای واکسینه از این جنبه دیده نشد. روی هم رفته نتایج بهدستآمده نشاندهنده مؤثر بودن سازواره ژنی طراحیشده بهعنوان یک کاندید واکسن مناسب در درمان التهابات ناشی از هلیکوباکتر پیلوری ازجمله کارسینوم معده میباشد.
سپاسگزاری
نویسندگان این مقاله از معاونت علمی و فناوری دانشگاه آزاد اسلامی واحد فلاورجان و همکاران محترم مرکز تحقیقات بیوتکنولوژی دانشگاه آزاد اسلامی واحد شهرکرد به دلیل همکاری صمیمانه در اجرای این پژوهش کمال امتنان را دارند. و مطالعه حاضر منتج از پایاننامه است که با کد 1724812835810611400162428555 در دانشگاه آزاد اسلامی واحد فلاورجان تصویب شده است.
حامی مالی: ندارد.
تعارض در منافع: وجود ندارد.
References:
1- Suarez G, Reyes VE, Beswick EJ. Immune Response to H Pylori. World J Gastroenterology: WJG 2006; 12(35): 5593-8.
2- Meliț LE, Mărginean CO, Săsăran MO. The Challenges of Eradicating Pediatric Helicobacter Pylori Infection in the Era of Probiotics. Children 2022; 9(6): 795.
3- Lee A, O'Rourke J, De Ungria MC, Robertson B, Daskalopoulos G, Dixon MF. A standardized mouse model of Helicobacter pylori infection: introducing the Sydney strain. Gastroenterology 1997; 112(4): 1386-97.
4- Correa P. Human Gastric Carcinogenesis: A Multistep And Multifactorial Process—First American Cancer Society Award Lecture On Cancer Epidemiology And Prevention. Cancer Res 1992; 52(24): 6735-40.
5- Verbeke H, Geboes K, Van Damme J, Struyf S. The Role of CXC Chemokines in the Transition of Chronic Inflammation to Esophageal and Gastric Cancer. Biochim Biophys Acta 2013; 1835(2): 117-29.
6- Backert S, Naumann M. What a disorder: proinflammatory signaling pathways induced by Helicobacter pylori. Trends Microbiol 2010; 18(11): 479-86.
7- Koosirirat C, Linpisarn S, Changsom D, Chawansuntati K, Wipasa J. Investigation of the Anti-Inflammatory Effect of Curcuma Longa in Helicobacter Pylori-Infected Patients. International immunopharmacology 2010; 10(7): 815-8.
8- Rad R, Brenner L, Krug A, Voland P, Mages J, Lang R, et al. Toll-like receptor–dependent activation of antigen-presenting cells affects adaptive immunity to Helicobacter pylori. Gastroenterology 2007; 133(1): 150-63. e3.
9- Bengmark S, Mesa MD, Gil A. Plant-Derived Health-The Effects of Turmeric and Curcuminoids. Nutricion Hospitalaria 2009; 24(3): 273-81.
10- Yang YJ, Chen PC, Lai FP, Tsai PJ, Sheu BS. Probiotics-Containing Yogurt Ingestion and H. pylori Eradication Can Restore Fecal Faecalibacterium prausnitzii Dysbiosis in H. pylori-Infected Children. Biomedicines 2020; 8(6): 146.
11- Shakhatreh MAK, Khabour OF, Alzoubi KH, Banihani MN, Abu-Siniyeh A, Bashir NA, et al. The Influence of IL-1B Gene Polymorphisms on H. Pylori Infection and Triple Treatment Response among Jordanian Population. Appl Clin Genet 2020; 13: 139-45.
12- Hu Y, Zhang M, Lu B, Dai J. Helicobacter pylori and Antibiotic Resistance, A Continuing and Intractable Problem. Helicobacter 2016; 21(5): 349-63.
13- Nejati S, Karkhah A, Darvish H, Validi M, Ebarhimpour S, Nouri HR. Influence of Helicobacter Pylori Virulence Factors Caga and Vaca on Pathogenesis of Gastrointestinal Disorders. Microb Pathog 2018; 1; 117: 43-8.
14- Wu D, Cao M, Peng J, Li N, Yi S, Song L, et al. The Effect of Trimethylamine N-Oxide on Helicobacter Pylori-Induced Changes of Immunoinflammatory Genes Expression in Gastric Epithelial Cells. Int Immunopharmacol 2017; 43: 172-8.
15- Roszczenko-Jasińska P, Wojtyś MI, Jagusztyn-Krynicka EK. Helicobacter Pylori Treatment in the Post-Antibiotics Era—Searching for New Drug Targets. Appl Microbiol Biotechnol 2020; 104(23): 9891-905.
16- El-Shouny WA, Ali SS, Hegazy HM, Abd Elnabi MK, Ali A, Sun J. Syzygium Aromaticum L.: Traditional Herbal Medicine Against Caga and Vaca Toxin Genes-Producing Drug Resistant Helicobacter Pylori. J Tradit Complement Med 2019; 10(4): 366-77.
17- Banga Ndzouboukou JL, Lei Q, Ullah N, Zhang Y, Hao L, Fan X. Helicobacter pylori adhesins: HpaA a potential antigen in experimental vaccines for H. pylori. Helicobacter 2021; 26(1): e12758.
18- Mirzaei N, Poursina F, Moghim S, Rashidi N, Ghasemian Safaei H. The Study of H. Pylori Putative Candidate Factors for Single- and Multi-Component Vaccine Development. Crit Rev Microbiol 2017; 43(5): 631-50.
19- Aliramaei MR, Khorasgani MR, Rahmani MR, Zarkesh Esfahani SH, Emamzadeh R. Expression of Helicobacter Pylori Cagl Gene In Lactococcus Lactis MG1363 and Evaluation of Its Immunogenicity as an Oral Vaccine in Mice. Microb Pathog 2020; 142: 103926.
20- Pan X, Ke H, Niu X, Li S, Lv J, Pan L. Protection Against Helicobacter Pylori Infection in BALB/C Mouse Model ذy Oral Administration of Multivalent Epitope-Based Vaccine of Cholera Toxin B Subunit-HUUC. Front Immunol 2018; 9: 1003.
21- Czinn SJ, Blanchard T. Vaccinating Against Helicobacter Pylori Infection. Nat Rev Gastroenterol Hepatol 2011; 8(3): 133-40.
22- Miyashita M, Joh T, Watanabe K, Todoroki I, Seno K, Ohara H, et al. Immune Responses in Mice to Intranasal and Intracutaneous Administration of a DNA Vaccine Encoding Helicobacter Pylori-Catalase. Vaccine 2002; 20(17-18): 2336-42.
23- Oñate AA, Céspedes S, Cabrera A, Rivers R, González A, Muñoz C, et al. A DNA Vaccine Encoding Cu,Zn Superoxide Dismutase Of Brucella Abortus Induces Protective Immunity In BALB/C Mice. Infect Immun 2003; 71(9): 4857-61.
24- Bai Y, Zhang YL, Chen Y, Jin JF, Zhang ZS, Zhou DY. Cloning and Expression and Immunogenicity of Helicobacter Pylori Baba2 Gene. World J Gastroenterol 2004; 10(17): 2560-2.
25- Cassataro J, Velikovsky CA, De La Barrera S, Estein SM, Bruno L, Bowden R, et al. A DNA vaccine coding for the Brucella outer membrane protein 31 confers protection against B. melitensis and B. Ovis Infection by Eliciting a Specific Cytotoxic Response. Infect Immun 2005; 73(10): 6537-46.
26- Doosti A, Ghasemi-Dehkordi P, Javadi GR, Sardari S, Shokrgozar MA. DNA Vaccine Encoding The Omp31 Gene Of Brucella Melitensis Induces Protective Immunity In BALB/C Mice. Res J Biol Sci 2009; 4(1): 126-31.
27- Zhao YX, Qi JH, Zhang H, Duan GC, Xi YL. Construction, Expression and Purification of Ureb-Omp11 Fusion Protein of Helicobacter Pylori and its Immunocompetence. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2007; 23(10): 906-10.
28- Al-Mariri A, Akel R, Abbady AQ. A DNA Vaccine Encoding P39 and Sp41 of Brucella Melitensis Induces Protective Immunity in BALB/C Mice. Arch Med Vet 2014; 46(1): 53-62.
نوع مطالعه: پژوهشي |
موضوع مقاله:
زیست شناسی
دریافت: 1401/3/25 | پذیرش: 1401/7/17 | انتشار: 1401/12/15
دریافت: 1401/3/25 | پذیرش: 1401/7/17 | انتشار: 1401/12/15
ارسال پیام به نویسنده مسئول
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |
