

دوره 29، شماره 2 - ( اردیبهشت 1400 )
جلد 29 شماره 2 صفحات 3533-3524 |
برگشت به فهرست نسخه ها


![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Hamid M, Shojaei Z. Analysis of Methylation Status in Promoter Region of Γ- Globin Gene in Carrier and Affected Β-Thalassemia Patients with High Level of Fetal Hemoglobin in Comparison with Normal Individuals. JSSU 2021; 29 (2) :3524-3533
URL: http://jssu.ssu.ac.ir/article-1-5104-fa.html
URL: http://jssu.ssu.ac.ir/article-1-5104-fa.html
حمید محمد، شجاعی زهره. بررسی الگوی متیلاسیون ناحیه پروموتری ژن گاماگلوبین در ناقلین و بیماران مبتلا به تالاسمی بتا با هموگلوبین جنینی بالا در مقایسه با افراد طبیعی. مجله علمي پژوهشي دانشگاه علوم پزشكي شهید صدوقی يزد. 1400; 29 (2) :3524-3533



متن کامل [PDF 1248 kb]
(545 دریافت)
| چکیده (HTML) (1315 مشاهده)
متن کامل: (1462 مشاهده)
مقدمه
هموگلوبین جنینی، نوعی هموگلوبین غالب در دوره جنینی است که تنها مدت کوتاهی بعد از تولد باقی میماند و زمانی تولید آن کاهش مییابد، که هموگلوبین بالغین رو به افزایش رود. مقادیر بالای هموگلوبین جنینی فواید سازمان یافته بالینی و گستردهای را در بیماران بتا تالاسمی و بیماری کم خونی سلولهای داسی شکل ایجاد میکند. افزایش هموگلوبین جنینی باعث افزایش هموگلوبین کل و کاهش علائم مربوط به تالاسمی میگردد. بیمارانی که هموگلوبین جنین بالاتری دارند عوارض بالینی خفیفتری را نشان میدهند. دلیل عمده آن این است که تشکیل هموگلوبین جنینی میتواند تا حدی تأثیرات مخرب تولید ناقص بتاگلوبین را جبران نماید (1). شناسایی ژنهای دخیل در ایجاد تنوع در تولید هموگلوبین جنینی و مکانیسمهای مولکولی کهبه وسیله آنها اهداف درمانی جدید شناسایی شود میتواند به توسعه داروهای جدید در این زمینه کمک کند. تغییرات اپی ژنتیکی که تا به امروز بهطور گسترده در انسان مورد مطالعه بوده، متیلاسیون سیتوزین DNA است. این شامل افزودن کووالانسی یک گروه متیل از دهنده متیل S -آدنوزیل متیونین به کربن موقعیت 1 سیتوزین در CpG توسط یک خانواده از آنزیمها به نام DNA methyltransferases (DNMTs) میشود (2). در حدود 30 میلیون جایگاه CpG در سراسر ژنوم وجود دارد که در آنها متیلاسیون صورت میگیرد. متیلاسیونDNA بهطور کلی با سرکوب رونویسی همراه است علاوه بر این با فرایندهای بسیار تخصصی مانند حکگذاری ژنومی و غیرفعالسازی کروموزوم X نیز در ارتباط است (3). چندین روش برای چگونگی عمل متیلاسیون DNA در خاموشی و مهار بیان ژن پیشنهاد شده است، دخالت مستقیم، اولین روش است که طی آن متیلاسیون بهطور مستقیم مانع نشستن فعالکنندههای رونویسی و فاکتورهای اتصال میشود. دومین روش پیشنهادی، وجود فاکتورهای سرکوبگر اختصاصی است. روش دیگر غیرفعال شدن ژن به دلیل تغییر وضعیت کروماتین از حالت باز و فعال به بسته و غیر فعال بهواسطه متیلاسیون DNA است (4). مطالعات نشان میدهد که متیلاسیون مناطق CpG در پروموتر نقش مهمی درسوئیچ ژن گاما به بتاگلوبین و در فعال شدن مجدد بیان گاماگلوبین توسط بازدارندههای DNA متیل ترانسفرازها ایفا میکند (5) در طی بیان ژنهای خوشه ژنی بتاگلوبین جایگاههای CpG در پروموتر ژنها غیرمتیله میباشند درحالیکه با خاموش شدن ژنها این جایگاهها تمایل به متیله شدن دارند (3). متیلاسیون CpG ممکن است مانعی برای تشکیل کمپلکس آغازگر نسخهبرداری بوده و بهطور غیرمستقیم از تغییر وضعیت کروماتین پیشگیری کند . شواهد نشان میدهد که متیلاسیون اثری روی تشکیل نوکلئوزوم ندارد اما DNA متیله شده پروتئینهای متصل شونده به متیل را فراخوانده که با هیستون دی استیلاز برهمکنش کرده و در تغییرات کروماتین نقش دارد (6). ناحیه CpG متیله ممکن است برای مهار رونویسی، از طریق دخالت مستقیم در اتصال فاکتور رونویسی و یا از طریق اتصال با پروتئینهای اس Stage Selector Protein (SSP) عمل کند و یا بهواسطه تغییر در ساختار کروماتین و فشرده شدن آن مانع رونویسی گردد (7). این حقیقت که فعال شدن ژن گاماگلوبین در سلولهای طبیعی تولیدکننده سلولهای خونی در ارتباط تنگاتنگ با هیپومتیلاسیون سایت CpG در ده جفت باز بالادست نقطه شروع رونویسی است، مشخص شده است این سایت محل اتصال وابسته به متیلاسیون فاکتورهای رونویسی خاصی از جمله کمپلکس پروتئینی SSP است (8). در مطالعات روی پروموتر ژنهای گاما و بتا گلوبین نیز مشخص شده است که میزان بیان آنها با متیلاسیون پروموترشان رابطه معکوس دارد به این معنا که در سلولهای خونی جنین پروموتر گاماگلوبین هیپومتیله است و در طی رشد جنین بیشتر و بیشتر متیله میشود تا جایی که در سلولهای خونی بالغین هایپرمتیله میشود (9). در این مطالعه میزان همراهی الگوی متیلاسیون پروموتر ژن گاماگلوبین در ناقلین و بیماران مبتلا به تالاسمی بتا با هموگلوبین جنینی بالا در مقایسه با افراد طبیعی مورد بررسی قرار گرفت.
روش بررسی
نمونهها: در مطالعه حاضر، که بهصورت مورد - شاهدی انجام گرفته است. شرکت کنندگان در این مطالعه (زن و مرد) با حداقل 2 سال و حداکثر 50 سال از مراجعهکنندگان به آزمایشگاه ژنتیک نرگس اهواز و انستیتو پاستور ایران انتخاب شدند. از نظر سن و جنس مورد و شاهد همسان انتخاب گردید. پس از اخذ رضایتنامه از افراد نرمال و بیماران تالاسمی، 30 بیمار تالاسمی ماژور و تالاسمی مینور همراه با هموگلوبین جنینی بالا (3/2 الی 98 درصد) و 30 فرد سالم بهعنوان افراد کنترل انتخاب شدند. در میان 30 نفر بررسی شده 15 نفر بتا تالاسمی مینور و 15 نفر بتا تالاسمی ماژور بودند. نمونهگیری بر اساس دادههای خونشناسی و الکتروفورز هموگلوبین از بین بیماران انجام شد. حجم نمونه با استفاده از روش مورگان تعداد 60 نمونه (30 نمونه افراد با هموگلوبین جنینی بالا (3/2 الی 98 درصد) و 30 نمونه شاهد) تعیین گردید. معیارهای ورود به مطالعه نمونهها افرادی بودند که هموگلوبین جنین آنها بالاتر 3/2 درصد و مبتلا یا ناقل تالاسمی بتاگلوبین بودند. معیارهای ورود به مطالعه در گروه شاهد شامل نمونههایی بودند که افراد فاقد هر گونه موتاسیون در ژن بتاگلوبین بوده و هموگلوبین جنینی آنها از 1 درصد پایینتر است. این مطالعه بهصورت کاملاً محرمانه و با رعایت کامل موازین، بهطوریکه مشخصات فردی نمونههای مورد مطالعه ناشناخته و با کد اختصاری علامتگذاری شده بودند.
استخراج DNA
نمونه خون در حدود دو میلیلیتر در لولههای حاوی ضدانعقاد EDTA جمعآوری گردید و استخراج DNA به روش Salting Out انجام گرفت. از دستگاه نانودراپ برای خواندن غلظت DNA بر حسب میکروگرم بر میلیلیتر استفاده گردید.
طراحی آغازگرها
برای تکثیر نواحی پروموتری ژنهای گاماگلوبین Gγ و گاما گلوبین Aγ در نمونههای جمعیت مورد مطالعه آغازگرهای پیشرو و معکوس با استفاده از نرمافزار Gene Runner version 4.0.9.2 beta و Methprimer و براساس توالی Acc. No: U01317.1 طراحی شد. در ابتدا بعد از بهدست آوردن توالی اصلی، به کمک Methprimer توالی کانورت شد. دی نوکلئوتیدهای CpG مشخص شد. آغازگرها طوری طراحی شدند که CpGهای حدود 300 نوکلئوتید در ناحیه بالادست و اگزون اول ژن گاماگلوبین را پوشش دهند. توالی آغازگرهای طراحی شده در جدول 1 آمده است.
تیمار سدیم بیسولفیت DNA، واکنش PCR و تعیین توالی
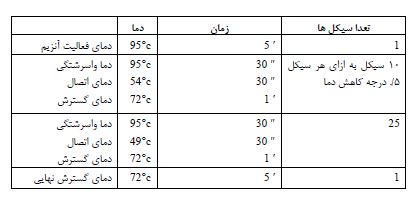
سدیم بیسولفیت میتواند سیتوزین را دآمینه یا کانورت نماید و به یوراسیل تبدیل کند، اما سیتوزین متیله را تحت تاثیر قرار نمیدهد. که بهمنظور انجام این روش از کیت EZ DNA Methylation-Lightning™ استفاده شد. سپس واکنش PCR جهت تکثیر نواحی انتخابی براساس برنامه جدول 2 انجام گردید. محصول PCR بر روی ژل آگارز 1/5% الکتروفورز شد. پس از اتمام الکتروفورز و اطمینان از عدم وجود باند غیر اختصاصی نمونهها در حجم μl 25 جهت انجام تعیین توالی ارسال شد. آغازگری که برای تعیین توالی استفاده شد همان آغازگر معکوسی بود که در تکثیر قطعات مورد مطالعه بهکار رفته بود.
تفسیر نمودار تعیین توالی
نمونه های تعیین توالی شده را با استفاده از نرمافزارهای Chromas وABI Sequence Scanner مورد بررسی قرار گرفتند. در این تحقیق ابتدا از برنامه Chromas برای بررسی اجمالی توالیها استفاده شد ، برای به دست آوردن ارتفاع پیکها جهت محاسبه درصد متیلاسیون از نرمافزار ABI Sequence Scanner استفاده شد. نسبت سایز پیکها متناسب با درصد کلی بازهای سیتوزینی است که متیله هستند بنابراین سطح متیلاسیون هر جایگاه CpG درون قطعه تکثیری از طریق فرمول (C/(C+T)*100) اندازهگیری شد (11, 10). درصد متیلاسیون کل قطعه تکثیری نیز با کمک میانگین درصد متیلاسیون همه جایگاهها بهدست آمد. لازم به ذکر است که فرمول فوق در زمان استفاده از پرایمر پیشرو کاربرد دارد و در صورتی که نمونهها با پرایمر معکوس تعیین توالی شده باشند باید از فرمول(G/(G+A)*100) استفاده کرد ( شکل1).
جدول 1: توالی آغازگرهای طراحی شده برای تکثیر نواحی تنظیمی ژن گاماگلوبین، سایز و محل قطعه تکثیری

جدول 2: برنامه Touch down PCR واکنش PCR برای تکثیر نواحی تنظیمی ژن گاماگلوبین

شکل 1: چگونگی انجام تجزیه و تحلیل دادههای تعیین توالی از یک جایگاه CpG با استفاده از آغازگر پیشرو پیک کروماتوگرام برای تیمین (نماینده سیتوزین غیرمتیله که در طول تیمار بیسولفیت به یوراسیل تبدیل شد) و سیتوزین (نماینده ی سیتوزین متیله که در طول تیمار بیسولفیت بدون تغییر باقی مانده است)
تجزیه وتحلیل آماری
جهت بررسی آماری نتایج بهدست آمده از آزمون آماری T- test با درجه اطمینان 95 درصد استفاده گردید و در آن درصدهای متیلاسیون هر جایگاه در گروه بیمار و نرمال با یکدیگر مقایسه گردید. بر این اساس مقدار p-value کمتر از 0/05 به معنای اختلاف معنیدار بین دو گروه انتخاب شد. دادهها به وسیله نرمافزار SPSS version 16 تجزیه و تحلیل شد.
ملاحظات اخلاقی
این تحقیق با گرنت و کد شماره 1033 و کد اخلاقی IR.PII.REC.1397.56 به تایید انستیتو پاستور ایران رسیده است.
نتایج
در این مطالعه (زن و مرد) با حداقل 2 سال و حداکثر 50 سال انتخاب شدند که از نظر سن و جنس مورد و شاهد همسان انتخاب شدند. از 60 بیمار و فرد سالم انتخاب شده در مطالعه 29 نفر خانم (48/33%) و 31 نفر آقا (51/67%) بودند (0/2=P). میانگین سنی بیماران و افراد شاهد به ترتیب 6±22 و 4±24 سال بود (0/5=P). پس از کانورت ژنوم افراد مورد بررسی، ناحیه پرموتری ژن بتاگلوبین تکثیر شده و بعد از الکتروفورز بر روی ژل آگارز قطعات 447 جفت بازی مشاهده شدند. سپس نمونهها برای بررسی شش جایگاه CpG برای هر نمونه شامل 162- ، 53- ، 50- ، 6+، 17+، 50+ تعیین توالی شدند. در صورتیکه در نتیجه تعیین توالی در موقعیتهای ذکر شده فقط یک پیک سیتوزین در جایگاه CG وجود داشته به این معناست که جایگاه CpG اولیه تماما متیله (100% متیله) بوده است .در صورتی که در این موقعیتها فقط پیک T باشد، به این معناست که جایگاه تماماً غیرمتیله) 0%متیله( بوده و در نهایت در صورتی که هر دو پیک C/T بالا آمده باشد وضعیت متیلاسیون نسبی گزارش میشود. برای توضیح بیشتر متیلاسیون نسبی میتوان گفت با حضور دو نسخه از هر کروموزوم در یک سلول، میتواند نسبتهای 0%، 50% و یا 100% متیله در یک CpG وجود داشته باشد. هنگام استخراجDNA ، یک ترکیبی ازDNA هزاران سلول تخلیص میشود بسته به اینکه چه مقدار از سلولها غیرمتیله، نیمه متیله یا تمام متیله باشند میتوانید یک سطح از متیلاسیون از 0 تا 100% را بهدست آورد از آنجا که تعیین توالی نمونههای تیمار شده با بیسولفیت دارای یک میزان خطا است در این تحقیق 80 تا 100% را بهعنوان تمام متیله 20 تا 80% به عنوان متیلاسیون نسبی انتخاب گردید و 40 تا 60% حالت نیمه متیله در نظر گرفته شد (12). نتایج بهدست آمده از بررسی توالیهای و تعیین درصد متیلاسیون افراد بیمار در جدول 3 آمده است که نتایج این جدول را میتوان بهصورت زیر خلاصه کرد: CpG ناحیه 162- در چهار نمونه 27 تا 30 تفاوت قابل توجهی در درصد متیلاسیون نشان داد بهطوریکه در سه نمونه که به ترتیب دارای درصد هموگلوبین جنینی 95/2،98 و98/3 بودند، حالت نیمه متیله دیده شد که تفاوت واضحی با نمونههای نرمال داشت (شکل 2) و یک نمونه نیز دارای وضعیت متیلاسیون نسبی با درصد متیلاسیون 75 بود. در CpG جایگاه 53- نیز وضعیت متیلاسیون نسبی با حدود متیلاسیون بین 62 تا 78 درصد در سه نمونه دیده شد که این نمونهها نیز دارای درصد هموگلوبین جنینی 18/8 ، 68 و 98/3 بودند. جایگاه 6+ نیز در شش نمونه وضعیت متیلاسیون نسبی با رنجی بین 61 الی 78 درصد نشان داد. جایگاههای 17+ و 50+ نیز هر کدام در یک نمونه وضعیت متیلاسیون نسبی نشان دادند. نتایج درصد متیلاسیون نمونههای نرمال در محدوده میانگین متیلاسیون بین5/89 الی 97 درصد بود و میتوان گفت تمام نمونهها تمام متیله بودند. اگرچه در یک نمونه نرمال CpG موقعیت 17+ دارای متیلاسیون 72% بود. مقدار p-value درجایگاههای 162- ، 53- و 6+ کمتر از 5 درصد بود و میتوان اینگونه نتیجه گرفت که اختلاف متیلاسیون در این جایگاهها بین دو گروه نرمال و بیمار معنیدار میباشد. این در حالی است که سه جایگاه دیگر شامل 50- ، 17+ و 50+ با توجه به مقدار p-value بیشتر از 5 درصد فاقد اختلاف معنیداری بین دو گروه مورد بررسی است. میانگین متیلاسیون کل در دو گروه بیمار و نرمال نیز دارای اختلاف قابل معنیداری بود(0/05>P).
جدول 3: درصد متیلاسیون، هموگلوبین جنینی و نوع موتاسیون ژن بتا گلوبین در نمونههای بیماران و ناقلین تالاسمی بتا
.JPG)

شکل 2: وضعیت نیمه متیله در جایگاه CpG 162- ژن گاما گلوبین
بحث
مطالعه حاضر بر روی افراد مبتلا به بتا تالاسمی ماژور یا مینور با هموگلوبین جنینی بالای3 درصد انجام گرفت و الگوی متیلاسیون پروموتر ژنهای گاماگلوبین این افراد با روش Bisulfite Sequencing مورد بررسی قرار گرفت. بهطور خلاصه، نتایج مطالعه حاضر نشان میدهد 21 نمونه از 30 نمونه بیماران و ناقلین بتا در تمام جایگاههای CpG مورد بررسی تمامأ متیله (81 الی 100%) بوده اند. 9 نمونه باقی مانده در حداقل یک سایتCpG وضعیت متیلاسیون نسبی یا نیمه متیله داشتهاند که حدود هموگلوبین جنینی این گروه از4/4 تا 98/3 درصد با میانگینی در حدود61 درصد متغیر بوده است. این در حالی است که تمام 30 نمونه نرمال بررسی شده تمامأ متیله بودهاند. از میان 9 نمونه ذکر شده در بالا که حداقل دارای یک جایگاه با وضعیت متیلاسیون متفاوت از نرمال بودهاند، 4 نمونه بهدلیل تعدد جایگاههای با وضعیت متیلاسیون نسبی دارای میانگین متیلاسیون زیر80 درصد ( 75/7 الی 79/2 درصد) نیز بودهاند، 3 نمونه این گروه دارای وضعیت نیمه متیله (40 الی 56 درصد) در CpG موقعیت 162- بودهاند. بر طبق آزمون آماری t-test محاسبه شده برای درصد متیلاسیون هر جایگاه در دو گروه نرمال و بیمار مشخص گردید که اختلاف متیلاسیون در 3 جایگاه 162- ،53- و 6+ میان این دو گروه معنیدار میباشد .(p <0.05) به بیان دیگر، احتمالأ اختلاف در میزان هموگلوبین جنینی در افراد بیمار و نرمال میتواند بهدلیل تفاوت در وضعیت متیلاسیون این سه جایگاه باشد. با این حال به این دلیل 21 نمونه از 30 نمونه مورد بررسی تفاوت قابل توجهی در سطح متیلاسیون نسبت به گروه نرمال نداشتهاند شاید بهتوان اینگونه نتیجه گرفت که دی متیلاسیون پروموتر ژن گاماگلوبین به خصوص سه جایگاه 162-، 53- و 6+ عامل ضروری در بیان مجدد ژن گاما گلوبین در بزرگسالی نباشد ولی احتمالأ عامل کمکی مهمی در این بیان مجدد است. جایگاه 6+ در محدوده اتصال آنزیمRNA Polymerase است. این امر میتواند دلیل گویایی بر معناداری اختلاف متیلاسیون این جایگاه در دو گروه مورد بررسی باشد. شاید متیلاسیون CpG جایگاه 6+ باعث کاهش میل آنزیم RNA Polymerase به اتصال در این محل گردد و از این طریق باعث کاهش بیان ژن گاماگلوبین و بهدنبال آن کاهش مقدار گاماگلوبین شود (13). جایگاه 53- بخشی از SSE است که محل اتصال پروتئینهای SSP و یا Sp1 در دورههای مختلف رشد است. جایگاه SSE یک توالی حفاظت شده در گونههایی است که در مرحله جنینی بیان ژن گاماگلوبین را دارند و نقش اساسی در تنظیم بیان ژن گاماگلوبین در طول رشد دارد. زمانی که جایگاه CpG دی متیله باشد اتصال پروتئین خاص اریتروئیدی SSP به جایگاه SSE صورت میگیرد و باعث فراخوانی Locus control region (LCR) به سمت پروموتر ژن گاماگلوبین میشود، از طرف دیگر متیلاسیون باعث القای اتصال ترجیحی پروتئین SP1 به SSE میشود، احتمال دادهاند که این اتصال بخشی از مکانیسم طبیعی سرکوب در سلولهای اریتروئیدی بزرگسالان است (17-14). ارتباط 53- در بیان هموگلوبین جنینی سالهاست که مشخص شده است بخشی از SSE است که بین نوکلئوتید 34 تا - 53 از پروموتر گاماگلوبین واقع شده است این عنصر در سوئیچ بیان ژن گاماگلوبین به بتاگلوبین در شرایط آزمایشگاهی دخالت میکند. SSE توسط SP1 و یا پروتئین SSP اشغال میگردد. کمپلکس SSP در فعالسازی گاماگلوبین نقش دارد در مقابل، اتصال SP1 به SSE بههمراه متیلاسیون جایگاه 53- باعث سرکوب گاماگلوبین میشود (17, 16). در خصوص جایگاه 162- میتوان گفت، با جستجوی حدود توالی این جایگاه در پایگاههای داده مشخص شد که این جایگاه پتانسیل اتصال فاکتور رونویسی MYB را دارا است. این پروتئین از جمله فاکتورهای رونویسی تاثیرگذار بر هماتوپویتیک است. در سلولهای اریتروئید اولیه که بیان بالای گاماگلوبین دارند سطح MYB پایین است و ارتباط معکوسی بین سطح این دو پروتئین در سلول وجود دارد، با این وجود از مکانیسم عمل این پروتئین و رابطه احتمالی آن با متیلاسیون پروموتر ژن گاماگلوبین اطلاعاتی وجود ندارد (18). هر چند پلیمورفیسمهایی در بالادست این ژن و در حد فاصل دو ژن MYB وHBS1L وجود دارد که در ارتباط با بقای هموگلوبین جنینی در بزرگسالی است و احتمال میدهند این ارتباط بهواسطه ژن MYB وجود داشته باشد (20, 19). بررسی الگوی متیلاسیون پروموتر ژن گاماگلوبین برای درک بهتر مکانیسم سوییچینگ گلوبین سالهاست که مورد بررسی است اما مطالعه در مورد وضعیت این جایگاه در افراد بتا تالاسمی با هموگلوبین جنینی بالا در سطح جهان معدود بوده است و بیشتر تمرکز روی ردههای سلولی بوده است. در تنها مطالعه مشابه انجام شده راجع به الگوی متیلاسیون پروموتر این ژن روی بیماران تالاسمی که در سال 2012 توسط Liu انجام شد، وضعیت متیلاسیون در چهار جایگاه CpG256-، 162-، 53- و 50- در ناحیه بالادست ژن گاماگلوبین در افراد مبتلا به بتاتالاسمی ماژور با هموگلوبین جنینی بالا در یکی از استانهای چین مورد بررسی قرارگرفتند. بر طبق این تحقیق تمام چهار جایگاه مورد بررسی چه در افراد نرمال و چه بیمار متیله بود ولی با این حال دو جایگاه 162- و 53- در افراد بیمار نسبت به نرمال هیپومتیله بود و در جایگاههای دیگر یعنی 256 – و 50- تفاوت معنیداری از نظر درجه متیلاسیون بین دو گروه مورد بررسی وجود نداشت. به طور کل میتوان گفت نتایج این تحقیق مشابه نتایج بهدست آمده در این تحقیق است (21). این مطالعه برای اولین بار روی نمونههای ایرانی نقش متیلاسیون DNA را مورد بررسی قرار میدهد.
نتیجهگیری
اگرچه اختلاف متیلاسیون در 3 جایگاه 162- ،53- و 6+ میان دو گروه بیمار و نرمال معنیدار میباشد اما به این دلیل که 21 نمونه از 30 نمونه مورد بررسی تفاوت قابل توجهی در سطح متیلاسیون نسبت به گروه نرمال نداشتهاند اینگونه میتوان نتیجه گرفت که دی متیلاسیون پروموتر ژن گاماگلوبین بهخصوص در سه جایگاه 162-، 53- و 6+ احتمالا نقش کمکی در بیان این ژن در دوران بزرگسالی دارد. همچنین بررسی فاکتورهای دیگر مرتبط با افزایش هموگلوبین جنینی از جمله عوامل تنظیمکننده سیس شامل پروموترهاPromoters ، تقویتکنندهها Enhacers، خاموشکنندهها Silencers، محدودکنندهها Insulators، نواحی تنظیمی تحت عنوان Locus-control region (LCR) و عوامل ترانس توصیه میگردد.
سپاسگزاری
از کلیه افراد شرکتکننده در این تحقیق و همکاران بخش پزشکی مولکولی انستیتو پاستور ایران تشکر و تقدیر بهعمل میآید. این طرح توسط انستیتو پاستور ایران با گرنت1033 حمایت مالی شده است.
حامی مالی: انستیتوپاستور ایران
تعارض منافع :وجود ندارد.
هموگلوبین جنینی، نوعی هموگلوبین غالب در دوره جنینی است که تنها مدت کوتاهی بعد از تولد باقی میماند و زمانی تولید آن کاهش مییابد، که هموگلوبین بالغین رو به افزایش رود. مقادیر بالای هموگلوبین جنینی فواید سازمان یافته بالینی و گستردهای را در بیماران بتا تالاسمی و بیماری کم خونی سلولهای داسی شکل ایجاد میکند. افزایش هموگلوبین جنینی باعث افزایش هموگلوبین کل و کاهش علائم مربوط به تالاسمی میگردد. بیمارانی که هموگلوبین جنین بالاتری دارند عوارض بالینی خفیفتری را نشان میدهند. دلیل عمده آن این است که تشکیل هموگلوبین جنینی میتواند تا حدی تأثیرات مخرب تولید ناقص بتاگلوبین را جبران نماید (1). شناسایی ژنهای دخیل در ایجاد تنوع در تولید هموگلوبین جنینی و مکانیسمهای مولکولی کهبه وسیله آنها اهداف درمانی جدید شناسایی شود میتواند به توسعه داروهای جدید در این زمینه کمک کند. تغییرات اپی ژنتیکی که تا به امروز بهطور گسترده در انسان مورد مطالعه بوده، متیلاسیون سیتوزین DNA است. این شامل افزودن کووالانسی یک گروه متیل از دهنده متیل S -آدنوزیل متیونین به کربن موقعیت 1 سیتوزین در CpG توسط یک خانواده از آنزیمها به نام DNA methyltransferases (DNMTs) میشود (2). در حدود 30 میلیون جایگاه CpG در سراسر ژنوم وجود دارد که در آنها متیلاسیون صورت میگیرد. متیلاسیونDNA بهطور کلی با سرکوب رونویسی همراه است علاوه بر این با فرایندهای بسیار تخصصی مانند حکگذاری ژنومی و غیرفعالسازی کروموزوم X نیز در ارتباط است (3). چندین روش برای چگونگی عمل متیلاسیون DNA در خاموشی و مهار بیان ژن پیشنهاد شده است، دخالت مستقیم، اولین روش است که طی آن متیلاسیون بهطور مستقیم مانع نشستن فعالکنندههای رونویسی و فاکتورهای اتصال میشود. دومین روش پیشنهادی، وجود فاکتورهای سرکوبگر اختصاصی است. روش دیگر غیرفعال شدن ژن به دلیل تغییر وضعیت کروماتین از حالت باز و فعال به بسته و غیر فعال بهواسطه متیلاسیون DNA است (4). مطالعات نشان میدهد که متیلاسیون مناطق CpG در پروموتر نقش مهمی درسوئیچ ژن گاما به بتاگلوبین و در فعال شدن مجدد بیان گاماگلوبین توسط بازدارندههای DNA متیل ترانسفرازها ایفا میکند (5) در طی بیان ژنهای خوشه ژنی بتاگلوبین جایگاههای CpG در پروموتر ژنها غیرمتیله میباشند درحالیکه با خاموش شدن ژنها این جایگاهها تمایل به متیله شدن دارند (3). متیلاسیون CpG ممکن است مانعی برای تشکیل کمپلکس آغازگر نسخهبرداری بوده و بهطور غیرمستقیم از تغییر وضعیت کروماتین پیشگیری کند . شواهد نشان میدهد که متیلاسیون اثری روی تشکیل نوکلئوزوم ندارد اما DNA متیله شده پروتئینهای متصل شونده به متیل را فراخوانده که با هیستون دی استیلاز برهمکنش کرده و در تغییرات کروماتین نقش دارد (6). ناحیه CpG متیله ممکن است برای مهار رونویسی، از طریق دخالت مستقیم در اتصال فاکتور رونویسی و یا از طریق اتصال با پروتئینهای اس Stage Selector Protein (SSP) عمل کند و یا بهواسطه تغییر در ساختار کروماتین و فشرده شدن آن مانع رونویسی گردد (7). این حقیقت که فعال شدن ژن گاماگلوبین در سلولهای طبیعی تولیدکننده سلولهای خونی در ارتباط تنگاتنگ با هیپومتیلاسیون سایت CpG در ده جفت باز بالادست نقطه شروع رونویسی است، مشخص شده است این سایت محل اتصال وابسته به متیلاسیون فاکتورهای رونویسی خاصی از جمله کمپلکس پروتئینی SSP است (8). در مطالعات روی پروموتر ژنهای گاما و بتا گلوبین نیز مشخص شده است که میزان بیان آنها با متیلاسیون پروموترشان رابطه معکوس دارد به این معنا که در سلولهای خونی جنین پروموتر گاماگلوبین هیپومتیله است و در طی رشد جنین بیشتر و بیشتر متیله میشود تا جایی که در سلولهای خونی بالغین هایپرمتیله میشود (9). در این مطالعه میزان همراهی الگوی متیلاسیون پروموتر ژن گاماگلوبین در ناقلین و بیماران مبتلا به تالاسمی بتا با هموگلوبین جنینی بالا در مقایسه با افراد طبیعی مورد بررسی قرار گرفت.
روش بررسی
نمونهها: در مطالعه حاضر، که بهصورت مورد - شاهدی انجام گرفته است. شرکت کنندگان در این مطالعه (زن و مرد) با حداقل 2 سال و حداکثر 50 سال از مراجعهکنندگان به آزمایشگاه ژنتیک نرگس اهواز و انستیتو پاستور ایران انتخاب شدند. از نظر سن و جنس مورد و شاهد همسان انتخاب گردید. پس از اخذ رضایتنامه از افراد نرمال و بیماران تالاسمی، 30 بیمار تالاسمی ماژور و تالاسمی مینور همراه با هموگلوبین جنینی بالا (3/2 الی 98 درصد) و 30 فرد سالم بهعنوان افراد کنترل انتخاب شدند. در میان 30 نفر بررسی شده 15 نفر بتا تالاسمی مینور و 15 نفر بتا تالاسمی ماژور بودند. نمونهگیری بر اساس دادههای خونشناسی و الکتروفورز هموگلوبین از بین بیماران انجام شد. حجم نمونه با استفاده از روش مورگان تعداد 60 نمونه (30 نمونه افراد با هموگلوبین جنینی بالا (3/2 الی 98 درصد) و 30 نمونه شاهد) تعیین گردید. معیارهای ورود به مطالعه نمونهها افرادی بودند که هموگلوبین جنین آنها بالاتر 3/2 درصد و مبتلا یا ناقل تالاسمی بتاگلوبین بودند. معیارهای ورود به مطالعه در گروه شاهد شامل نمونههایی بودند که افراد فاقد هر گونه موتاسیون در ژن بتاگلوبین بوده و هموگلوبین جنینی آنها از 1 درصد پایینتر است. این مطالعه بهصورت کاملاً محرمانه و با رعایت کامل موازین، بهطوریکه مشخصات فردی نمونههای مورد مطالعه ناشناخته و با کد اختصاری علامتگذاری شده بودند.
استخراج DNA
نمونه خون در حدود دو میلیلیتر در لولههای حاوی ضدانعقاد EDTA جمعآوری گردید و استخراج DNA به روش Salting Out انجام گرفت. از دستگاه نانودراپ برای خواندن غلظت DNA بر حسب میکروگرم بر میلیلیتر استفاده گردید.
طراحی آغازگرها
برای تکثیر نواحی پروموتری ژنهای گاماگلوبین Gγ و گاما گلوبین Aγ در نمونههای جمعیت مورد مطالعه آغازگرهای پیشرو و معکوس با استفاده از نرمافزار Gene Runner version 4.0.9.2 beta و Methprimer و براساس توالی Acc. No: U01317.1 طراحی شد. در ابتدا بعد از بهدست آوردن توالی اصلی، به کمک Methprimer توالی کانورت شد. دی نوکلئوتیدهای CpG مشخص شد. آغازگرها طوری طراحی شدند که CpGهای حدود 300 نوکلئوتید در ناحیه بالادست و اگزون اول ژن گاماگلوبین را پوشش دهند. توالی آغازگرهای طراحی شده در جدول 1 آمده است.
تیمار سدیم بیسولفیت DNA، واکنش PCR و تعیین توالی
سدیم بیسولفیت میتواند سیتوزین را دآمینه یا کانورت نماید و به یوراسیل تبدیل کند، اما سیتوزین متیله را تحت تاثیر قرار نمیدهد. که بهمنظور انجام این روش از کیت EZ DNA Methylation-Lightning™ استفاده شد. سپس واکنش PCR جهت تکثیر نواحی انتخابی براساس برنامه جدول 2 انجام گردید. محصول PCR بر روی ژل آگارز 1/5% الکتروفورز شد. پس از اتمام الکتروفورز و اطمینان از عدم وجود باند غیر اختصاصی نمونهها در حجم μl 25 جهت انجام تعیین توالی ارسال شد. آغازگری که برای تعیین توالی استفاده شد همان آغازگر معکوسی بود که در تکثیر قطعات مورد مطالعه بهکار رفته بود.
تفسیر نمودار تعیین توالی
نمونه های تعیین توالی شده را با استفاده از نرمافزارهای Chromas وABI Sequence Scanner مورد بررسی قرار گرفتند. در این تحقیق ابتدا از برنامه Chromas برای بررسی اجمالی توالیها استفاده شد ، برای به دست آوردن ارتفاع پیکها جهت محاسبه درصد متیلاسیون از نرمافزار ABI Sequence Scanner استفاده شد. نسبت سایز پیکها متناسب با درصد کلی بازهای سیتوزینی است که متیله هستند بنابراین سطح متیلاسیون هر جایگاه CpG درون قطعه تکثیری از طریق فرمول (C/(C+T)*100) اندازهگیری شد (11, 10). درصد متیلاسیون کل قطعه تکثیری نیز با کمک میانگین درصد متیلاسیون همه جایگاهها بهدست آمد. لازم به ذکر است که فرمول فوق در زمان استفاده از پرایمر پیشرو کاربرد دارد و در صورتی که نمونهها با پرایمر معکوس تعیین توالی شده باشند باید از فرمول(G/(G+A)*100) استفاده کرد ( شکل1).
جدول 1: توالی آغازگرهای طراحی شده برای تکثیر نواحی تنظیمی ژن گاماگلوبین، سایز و محل قطعه تکثیری
جدول 2: برنامه Touch down PCR واکنش PCR برای تکثیر نواحی تنظیمی ژن گاماگلوبین
شکل 1: چگونگی انجام تجزیه و تحلیل دادههای تعیین توالی از یک جایگاه CpG با استفاده از آغازگر پیشرو پیک کروماتوگرام برای تیمین (نماینده سیتوزین غیرمتیله که در طول تیمار بیسولفیت به یوراسیل تبدیل شد) و سیتوزین (نماینده ی سیتوزین متیله که در طول تیمار بیسولفیت بدون تغییر باقی مانده است)
تجزیه وتحلیل آماری
جهت بررسی آماری نتایج بهدست آمده از آزمون آماری T- test با درجه اطمینان 95 درصد استفاده گردید و در آن درصدهای متیلاسیون هر جایگاه در گروه بیمار و نرمال با یکدیگر مقایسه گردید. بر این اساس مقدار p-value کمتر از 0/05 به معنای اختلاف معنیدار بین دو گروه انتخاب شد. دادهها به وسیله نرمافزار SPSS version 16 تجزیه و تحلیل شد.
ملاحظات اخلاقی
این تحقیق با گرنت و کد شماره 1033 و کد اخلاقی IR.PII.REC.1397.56 به تایید انستیتو پاستور ایران رسیده است.
نتایج
در این مطالعه (زن و مرد) با حداقل 2 سال و حداکثر 50 سال انتخاب شدند که از نظر سن و جنس مورد و شاهد همسان انتخاب شدند. از 60 بیمار و فرد سالم انتخاب شده در مطالعه 29 نفر خانم (48/33%) و 31 نفر آقا (51/67%) بودند (0/2=P). میانگین سنی بیماران و افراد شاهد به ترتیب 6±22 و 4±24 سال بود (0/5=P). پس از کانورت ژنوم افراد مورد بررسی، ناحیه پرموتری ژن بتاگلوبین تکثیر شده و بعد از الکتروفورز بر روی ژل آگارز قطعات 447 جفت بازی مشاهده شدند. سپس نمونهها برای بررسی شش جایگاه CpG برای هر نمونه شامل 162- ، 53- ، 50- ، 6+، 17+، 50+ تعیین توالی شدند. در صورتیکه در نتیجه تعیین توالی در موقعیتهای ذکر شده فقط یک پیک سیتوزین در جایگاه CG وجود داشته به این معناست که جایگاه CpG اولیه تماما متیله (100% متیله) بوده است .در صورتی که در این موقعیتها فقط پیک T باشد، به این معناست که جایگاه تماماً غیرمتیله) 0%متیله( بوده و در نهایت در صورتی که هر دو پیک C/T بالا آمده باشد وضعیت متیلاسیون نسبی گزارش میشود. برای توضیح بیشتر متیلاسیون نسبی میتوان گفت با حضور دو نسخه از هر کروموزوم در یک سلول، میتواند نسبتهای 0%، 50% و یا 100% متیله در یک CpG وجود داشته باشد. هنگام استخراجDNA ، یک ترکیبی ازDNA هزاران سلول تخلیص میشود بسته به اینکه چه مقدار از سلولها غیرمتیله، نیمه متیله یا تمام متیله باشند میتوانید یک سطح از متیلاسیون از 0 تا 100% را بهدست آورد از آنجا که تعیین توالی نمونههای تیمار شده با بیسولفیت دارای یک میزان خطا است در این تحقیق 80 تا 100% را بهعنوان تمام متیله 20 تا 80% به عنوان متیلاسیون نسبی انتخاب گردید و 40 تا 60% حالت نیمه متیله در نظر گرفته شد (12). نتایج بهدست آمده از بررسی توالیهای و تعیین درصد متیلاسیون افراد بیمار در جدول 3 آمده است که نتایج این جدول را میتوان بهصورت زیر خلاصه کرد: CpG ناحیه 162- در چهار نمونه 27 تا 30 تفاوت قابل توجهی در درصد متیلاسیون نشان داد بهطوریکه در سه نمونه که به ترتیب دارای درصد هموگلوبین جنینی 95/2،98 و98/3 بودند، حالت نیمه متیله دیده شد که تفاوت واضحی با نمونههای نرمال داشت (شکل 2) و یک نمونه نیز دارای وضعیت متیلاسیون نسبی با درصد متیلاسیون 75 بود. در CpG جایگاه 53- نیز وضعیت متیلاسیون نسبی با حدود متیلاسیون بین 62 تا 78 درصد در سه نمونه دیده شد که این نمونهها نیز دارای درصد هموگلوبین جنینی 18/8 ، 68 و 98/3 بودند. جایگاه 6+ نیز در شش نمونه وضعیت متیلاسیون نسبی با رنجی بین 61 الی 78 درصد نشان داد. جایگاههای 17+ و 50+ نیز هر کدام در یک نمونه وضعیت متیلاسیون نسبی نشان دادند. نتایج درصد متیلاسیون نمونههای نرمال در محدوده میانگین متیلاسیون بین5/89 الی 97 درصد بود و میتوان گفت تمام نمونهها تمام متیله بودند. اگرچه در یک نمونه نرمال CpG موقعیت 17+ دارای متیلاسیون 72% بود. مقدار p-value درجایگاههای 162- ، 53- و 6+ کمتر از 5 درصد بود و میتوان اینگونه نتیجه گرفت که اختلاف متیلاسیون در این جایگاهها بین دو گروه نرمال و بیمار معنیدار میباشد. این در حالی است که سه جایگاه دیگر شامل 50- ، 17+ و 50+ با توجه به مقدار p-value بیشتر از 5 درصد فاقد اختلاف معنیداری بین دو گروه مورد بررسی است. میانگین متیلاسیون کل در دو گروه بیمار و نرمال نیز دارای اختلاف قابل معنیداری بود(0/05>P).
جدول 3: درصد متیلاسیون، هموگلوبین جنینی و نوع موتاسیون ژن بتا گلوبین در نمونههای بیماران و ناقلین تالاسمی بتا
شکل 2: وضعیت نیمه متیله در جایگاه CpG 162- ژن گاما گلوبین
بحث
مطالعه حاضر بر روی افراد مبتلا به بتا تالاسمی ماژور یا مینور با هموگلوبین جنینی بالای3 درصد انجام گرفت و الگوی متیلاسیون پروموتر ژنهای گاماگلوبین این افراد با روش Bisulfite Sequencing مورد بررسی قرار گرفت. بهطور خلاصه، نتایج مطالعه حاضر نشان میدهد 21 نمونه از 30 نمونه بیماران و ناقلین بتا در تمام جایگاههای CpG مورد بررسی تمامأ متیله (81 الی 100%) بوده اند. 9 نمونه باقی مانده در حداقل یک سایتCpG وضعیت متیلاسیون نسبی یا نیمه متیله داشتهاند که حدود هموگلوبین جنینی این گروه از4/4 تا 98/3 درصد با میانگینی در حدود61 درصد متغیر بوده است. این در حالی است که تمام 30 نمونه نرمال بررسی شده تمامأ متیله بودهاند. از میان 9 نمونه ذکر شده در بالا که حداقل دارای یک جایگاه با وضعیت متیلاسیون متفاوت از نرمال بودهاند، 4 نمونه بهدلیل تعدد جایگاههای با وضعیت متیلاسیون نسبی دارای میانگین متیلاسیون زیر80 درصد ( 75/7 الی 79/2 درصد) نیز بودهاند، 3 نمونه این گروه دارای وضعیت نیمه متیله (40 الی 56 درصد) در CpG موقعیت 162- بودهاند. بر طبق آزمون آماری t-test محاسبه شده برای درصد متیلاسیون هر جایگاه در دو گروه نرمال و بیمار مشخص گردید که اختلاف متیلاسیون در 3 جایگاه 162- ،53- و 6+ میان این دو گروه معنیدار میباشد .(p <0.05) به بیان دیگر، احتمالأ اختلاف در میزان هموگلوبین جنینی در افراد بیمار و نرمال میتواند بهدلیل تفاوت در وضعیت متیلاسیون این سه جایگاه باشد. با این حال به این دلیل 21 نمونه از 30 نمونه مورد بررسی تفاوت قابل توجهی در سطح متیلاسیون نسبت به گروه نرمال نداشتهاند شاید بهتوان اینگونه نتیجه گرفت که دی متیلاسیون پروموتر ژن گاماگلوبین به خصوص سه جایگاه 162-، 53- و 6+ عامل ضروری در بیان مجدد ژن گاما گلوبین در بزرگسالی نباشد ولی احتمالأ عامل کمکی مهمی در این بیان مجدد است. جایگاه 6+ در محدوده اتصال آنزیمRNA Polymerase است. این امر میتواند دلیل گویایی بر معناداری اختلاف متیلاسیون این جایگاه در دو گروه مورد بررسی باشد. شاید متیلاسیون CpG جایگاه 6+ باعث کاهش میل آنزیم RNA Polymerase به اتصال در این محل گردد و از این طریق باعث کاهش بیان ژن گاماگلوبین و بهدنبال آن کاهش مقدار گاماگلوبین شود (13). جایگاه 53- بخشی از SSE است که محل اتصال پروتئینهای SSP و یا Sp1 در دورههای مختلف رشد است. جایگاه SSE یک توالی حفاظت شده در گونههایی است که در مرحله جنینی بیان ژن گاماگلوبین را دارند و نقش اساسی در تنظیم بیان ژن گاماگلوبین در طول رشد دارد. زمانی که جایگاه CpG دی متیله باشد اتصال پروتئین خاص اریتروئیدی SSP به جایگاه SSE صورت میگیرد و باعث فراخوانی Locus control region (LCR) به سمت پروموتر ژن گاماگلوبین میشود، از طرف دیگر متیلاسیون باعث القای اتصال ترجیحی پروتئین SP1 به SSE میشود، احتمال دادهاند که این اتصال بخشی از مکانیسم طبیعی سرکوب در سلولهای اریتروئیدی بزرگسالان است (17-14). ارتباط 53- در بیان هموگلوبین جنینی سالهاست که مشخص شده است بخشی از SSE است که بین نوکلئوتید 34 تا - 53 از پروموتر گاماگلوبین واقع شده است این عنصر در سوئیچ بیان ژن گاماگلوبین به بتاگلوبین در شرایط آزمایشگاهی دخالت میکند. SSE توسط SP1 و یا پروتئین SSP اشغال میگردد. کمپلکس SSP در فعالسازی گاماگلوبین نقش دارد در مقابل، اتصال SP1 به SSE بههمراه متیلاسیون جایگاه 53- باعث سرکوب گاماگلوبین میشود (17, 16). در خصوص جایگاه 162- میتوان گفت، با جستجوی حدود توالی این جایگاه در پایگاههای داده مشخص شد که این جایگاه پتانسیل اتصال فاکتور رونویسی MYB را دارا است. این پروتئین از جمله فاکتورهای رونویسی تاثیرگذار بر هماتوپویتیک است. در سلولهای اریتروئید اولیه که بیان بالای گاماگلوبین دارند سطح MYB پایین است و ارتباط معکوسی بین سطح این دو پروتئین در سلول وجود دارد، با این وجود از مکانیسم عمل این پروتئین و رابطه احتمالی آن با متیلاسیون پروموتر ژن گاماگلوبین اطلاعاتی وجود ندارد (18). هر چند پلیمورفیسمهایی در بالادست این ژن و در حد فاصل دو ژن MYB وHBS1L وجود دارد که در ارتباط با بقای هموگلوبین جنینی در بزرگسالی است و احتمال میدهند این ارتباط بهواسطه ژن MYB وجود داشته باشد (20, 19). بررسی الگوی متیلاسیون پروموتر ژن گاماگلوبین برای درک بهتر مکانیسم سوییچینگ گلوبین سالهاست که مورد بررسی است اما مطالعه در مورد وضعیت این جایگاه در افراد بتا تالاسمی با هموگلوبین جنینی بالا در سطح جهان معدود بوده است و بیشتر تمرکز روی ردههای سلولی بوده است. در تنها مطالعه مشابه انجام شده راجع به الگوی متیلاسیون پروموتر این ژن روی بیماران تالاسمی که در سال 2012 توسط Liu انجام شد، وضعیت متیلاسیون در چهار جایگاه CpG256-، 162-، 53- و 50- در ناحیه بالادست ژن گاماگلوبین در افراد مبتلا به بتاتالاسمی ماژور با هموگلوبین جنینی بالا در یکی از استانهای چین مورد بررسی قرارگرفتند. بر طبق این تحقیق تمام چهار جایگاه مورد بررسی چه در افراد نرمال و چه بیمار متیله بود ولی با این حال دو جایگاه 162- و 53- در افراد بیمار نسبت به نرمال هیپومتیله بود و در جایگاههای دیگر یعنی 256 – و 50- تفاوت معنیداری از نظر درجه متیلاسیون بین دو گروه مورد بررسی وجود نداشت. به طور کل میتوان گفت نتایج این تحقیق مشابه نتایج بهدست آمده در این تحقیق است (21). این مطالعه برای اولین بار روی نمونههای ایرانی نقش متیلاسیون DNA را مورد بررسی قرار میدهد.
نتیجهگیری
اگرچه اختلاف متیلاسیون در 3 جایگاه 162- ،53- و 6+ میان دو گروه بیمار و نرمال معنیدار میباشد اما به این دلیل که 21 نمونه از 30 نمونه مورد بررسی تفاوت قابل توجهی در سطح متیلاسیون نسبت به گروه نرمال نداشتهاند اینگونه میتوان نتیجه گرفت که دی متیلاسیون پروموتر ژن گاماگلوبین بهخصوص در سه جایگاه 162-، 53- و 6+ احتمالا نقش کمکی در بیان این ژن در دوران بزرگسالی دارد. همچنین بررسی فاکتورهای دیگر مرتبط با افزایش هموگلوبین جنینی از جمله عوامل تنظیمکننده سیس شامل پروموترهاPromoters ، تقویتکنندهها Enhacers، خاموشکنندهها Silencers، محدودکنندهها Insulators، نواحی تنظیمی تحت عنوان Locus-control region (LCR) و عوامل ترانس توصیه میگردد.
سپاسگزاری
از کلیه افراد شرکتکننده در این تحقیق و همکاران بخش پزشکی مولکولی انستیتو پاستور ایران تشکر و تقدیر بهعمل میآید. این طرح توسط انستیتو پاستور ایران با گرنت1033 حمایت مالی شده است.
حامی مالی: انستیتوپاستور ایران
تعارض منافع :وجود ندارد.
References:
1- Hamid M, Mahjoubi F, Akbari MT, Arab A, Zeinali S, Karimipoor M. Molecular Analysis of Gamma-Globin Promoters, HS-111 and 3'HS1, In Beta-Thalassemia Intermedia Patients Associated with High Levels of Hb F. Hemoglobin 2009; 33(6): 428-38.
2- Rhee I, Bachman KE, Park BH, Jair KW, Yen RW, Schuebel KE, et al. DNMT1 and DNMT3b Cooperate to Silence Genes in Human Cancer Cells. Nature 2002; 416(6880): 552-6.
3- Jiang YH, Bressler J, Beaudet AL. Epigenetics and Human Disease. Annu Rev Genomics Hum Genet 2004; 5: 479-510.
4- Singal R, Ginder GD. DNA Methylation. Blood 1999; 93(12): 4059-70.
5- Kiefer CM, Hou C, Little JA, Dean A. Epigenetics of Beta-Globin Gene Regulation. Mutat Res 2008; 647(1-2): 68-76.
6- Hardison R. Hemoglobins from Bacteria to Man: Evolution of Different Patterns of Gene Expression. J Exp Biol 1998; 201: 1099-117.
7- Lavelle D, Vaitkus K, Hankewych M, Singh M, DeSimone J. Developmental Changes in DNA Methylation and Covalent Histone Modifications of Chromatin Associated with the Epsilon- Gamma- And Beta-Globin Gene Promoters in Papio Anubis. Blood Cells Mol Dis 2006; 36(2): 269-78.
8- Sengupta PK, Lavelle D, DeSimone J. Increased Binding of Sp1 to the Gamma-Globin Gene Promoter Upon Site-Specific Cytosine Methylation. Am J Hematol 1994; 46(3): 169-72.
9- Musallam KM, Taher AT, Cappellini MD, Sankaran VG. Clinical Experience with Fetal Hemoglobin Induction Therapy in Patients with Beta-Thalassemia. Blood 2013; 121(12): 2199-212.
10- Jiang M, Zhang Y, Fei J, Chang X, Fan W, Qian X, et al. Rapid Quantification of DNA Methylation by Measuring Relative Peak Heights in Direct Bisulfite-PCR Sequencing Traces. Lab Invest 2010; 90(2): 282-90.
11- Parrish RR, Day JJ, Lubin FD. Direct Bisulfite Sequencing for Examination of DNA Methylation with Gene and Nucleotide Resolution from Brain Tissues. Curr Protoc Neurosci 2012; Chapter 7: Unit 7 24.
12- Smallwood SA, Lee HJ, Angermueller C, Krueger F, Saadeh H, Peat J, et al. Single-Cell Genome-Wide Bisulfite Sequencing for Assessing Epigenetic Heterogeneity. Nat Methods 2014; 11(8): 817-20.
13- Makala L, Torres C, EL EC, Neunert C, Betty S. Pace. Fetal Hemoglobin Induction to Treat b-Hemoglobinopathies from Bench to Bedside. J Hematol Transfus 2014; 2(2): 1018.
14- Jane SM, Ney PA, Vanin EF, Gumucio DL, Nienhuis AW. Identification of a Stage Selector Element in the Human Gamma‐Globin Gene Promoter that Fosters Preferential Interaction with the 5′ HS2 Enhancer when in Competition with the Beta‐Promoter. EMBO J 1992; 11(8): 2961-9.
15- Gumucio D, Heilstedt-Williamson H, Gray T, Tarle S, Shelton D, Tagle D, et al. Phylogenetic Footprinting Reveals a Nuclear Protein which Binds to Silencer Sequences in the Human Gamma and Epsilon Globin Genes. Mol Cell Biol 1992; 12(11): 4919-29.
16- Sengupta PK, Lavelle D, Desimone J. Increased Binding of SP1 to the Γ‐Globin Gene Promoter Upon Site‐Specific Cytosine Methylation. Am J Hematol 1994; 46(3): 169-72.
17- Jane SM, Gumucio D, Ney P, Cunningham J, Nienhuis A. Methylation-Enhanced Binding of Sp1 to the Stage Selector Element of the Human Gamma-Globin Gene Promoter May Regulate Development Specificity of Expression. Mol Cell Biol 1993; 13(6): 3272-81.
18- Jiang J, Best S, Menzel S, Silver N, Lai MI, Surdulescu GL, et al. CMYB is Involved in the Regulation of Fetal Hemoglobin Production in Adults. Blood 2006; 108(3): 1077-83.
19- Thein SL, Menzel S, Peng X, Best S, Jiang J, Close J, et al. Intergenic Variants of HBS1L-MYB are Responsible for a Major Quantitative Trait Locus on Chromosome 6q23 Influencing Fetal Hemoglobin Levels in Adults. Proc Natl Acad Sci 2007; 104(27): 11346-51.
20- George OL, Ness SA. Situational Awareness: Regulation of the Myb Transcription Factor in Differentiation, The Cell Cycle and Oncogenesis. Cancers 2014; 6(4): 2049-71.
21- Liu Y, Cheng P, Liu Z, Liu R, Lai Y. Methylation Status of Γ-Globin Gene Promoter in Β-Thalassemia Major. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2012; 20(3): 642-5.
1- Hamid M, Mahjoubi F, Akbari MT, Arab A, Zeinali S, Karimipoor M. Molecular Analysis of Gamma-Globin Promoters, HS-111 and 3'HS1, In Beta-Thalassemia Intermedia Patients Associated with High Levels of Hb F. Hemoglobin 2009; 33(6): 428-38.
2- Rhee I, Bachman KE, Park BH, Jair KW, Yen RW, Schuebel KE, et al. DNMT1 and DNMT3b Cooperate to Silence Genes in Human Cancer Cells. Nature 2002; 416(6880): 552-6.
3- Jiang YH, Bressler J, Beaudet AL. Epigenetics and Human Disease. Annu Rev Genomics Hum Genet 2004; 5: 479-510.
4- Singal R, Ginder GD. DNA Methylation. Blood 1999; 93(12): 4059-70.
5- Kiefer CM, Hou C, Little JA, Dean A. Epigenetics of Beta-Globin Gene Regulation. Mutat Res 2008; 647(1-2): 68-76.
6- Hardison R. Hemoglobins from Bacteria to Man: Evolution of Different Patterns of Gene Expression. J Exp Biol 1998; 201: 1099-117.
7- Lavelle D, Vaitkus K, Hankewych M, Singh M, DeSimone J. Developmental Changes in DNA Methylation and Covalent Histone Modifications of Chromatin Associated with the Epsilon- Gamma- And Beta-Globin Gene Promoters in Papio Anubis. Blood Cells Mol Dis 2006; 36(2): 269-78.
8- Sengupta PK, Lavelle D, DeSimone J. Increased Binding of Sp1 to the Gamma-Globin Gene Promoter Upon Site-Specific Cytosine Methylation. Am J Hematol 1994; 46(3): 169-72.
9- Musallam KM, Taher AT, Cappellini MD, Sankaran VG. Clinical Experience with Fetal Hemoglobin Induction Therapy in Patients with Beta-Thalassemia. Blood 2013; 121(12): 2199-212.
10- Jiang M, Zhang Y, Fei J, Chang X, Fan W, Qian X, et al. Rapid Quantification of DNA Methylation by Measuring Relative Peak Heights in Direct Bisulfite-PCR Sequencing Traces. Lab Invest 2010; 90(2): 282-90.
11- Parrish RR, Day JJ, Lubin FD. Direct Bisulfite Sequencing for Examination of DNA Methylation with Gene and Nucleotide Resolution from Brain Tissues. Curr Protoc Neurosci 2012; Chapter 7: Unit 7 24.
12- Smallwood SA, Lee HJ, Angermueller C, Krueger F, Saadeh H, Peat J, et al. Single-Cell Genome-Wide Bisulfite Sequencing for Assessing Epigenetic Heterogeneity. Nat Methods 2014; 11(8): 817-20.
13- Makala L, Torres C, EL EC, Neunert C, Betty S. Pace. Fetal Hemoglobin Induction to Treat b-Hemoglobinopathies from Bench to Bedside. J Hematol Transfus 2014; 2(2): 1018.
14- Jane SM, Ney PA, Vanin EF, Gumucio DL, Nienhuis AW. Identification of a Stage Selector Element in the Human Gamma‐Globin Gene Promoter that Fosters Preferential Interaction with the 5′ HS2 Enhancer when in Competition with the Beta‐Promoter. EMBO J 1992; 11(8): 2961-9.
15- Gumucio D, Heilstedt-Williamson H, Gray T, Tarle S, Shelton D, Tagle D, et al. Phylogenetic Footprinting Reveals a Nuclear Protein which Binds to Silencer Sequences in the Human Gamma and Epsilon Globin Genes. Mol Cell Biol 1992; 12(11): 4919-29.
16- Sengupta PK, Lavelle D, Desimone J. Increased Binding of SP1 to the Γ‐Globin Gene Promoter Upon Site‐Specific Cytosine Methylation. Am J Hematol 1994; 46(3): 169-72.
17- Jane SM, Gumucio D, Ney P, Cunningham J, Nienhuis A. Methylation-Enhanced Binding of Sp1 to the Stage Selector Element of the Human Gamma-Globin Gene Promoter May Regulate Development Specificity of Expression. Mol Cell Biol 1993; 13(6): 3272-81.
18- Jiang J, Best S, Menzel S, Silver N, Lai MI, Surdulescu GL, et al. CMYB is Involved in the Regulation of Fetal Hemoglobin Production in Adults. Blood 2006; 108(3): 1077-83.
19- Thein SL, Menzel S, Peng X, Best S, Jiang J, Close J, et al. Intergenic Variants of HBS1L-MYB are Responsible for a Major Quantitative Trait Locus on Chromosome 6q23 Influencing Fetal Hemoglobin Levels in Adults. Proc Natl Acad Sci 2007; 104(27): 11346-51.
20- George OL, Ness SA. Situational Awareness: Regulation of the Myb Transcription Factor in Differentiation, The Cell Cycle and Oncogenesis. Cancers 2014; 6(4): 2049-71.
21- Liu Y, Cheng P, Liu Z, Liu R, Lai Y. Methylation Status of Γ-Globin Gene Promoter in Β-Thalassemia Major. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2012; 20(3): 642-5.
ارسال پیام به نویسنده مسئول
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |
