

دوره 30، شماره 12 - ( اسفند 1401 )
جلد 30 شماره 12 صفحات 6198-6185 |
برگشت به فهرست نسخه ها


Ethics code: IR.SUMS.REHAB.REC.1400.027
![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Faramarziyan N, Salesi M, Rezaei R, Kooshki Jahromi M. Effects of Six Weeks High-Intensity Interval Training on TGF-b1 and SMAD7 Genes Expression in the Kidney Tissue of Elderly Diabetic Male Wistar Rats. JSSU 2023; 30 (12) :6185-6198
URL: http://jssu.ssu.ac.ir/article-1-5825-fa.html
URL: http://jssu.ssu.ac.ir/article-1-5825-fa.html
فرامرزیان نرگس، ثالثی محسن، رضایی رسول، کوشکی جهرمی مریم. تاثیر شش هفته تمرین تناوبی شدید بر بیان ژنهای TGF-β1 و SMAD7 در بافت کلیه موشهای صحرایینر نژاد ویستار دیابتی سالمند. مجله علمي پژوهشي دانشگاه علوم پزشكي شهید صدوقی يزد. 1401; 30 (12) :6185-6198



متن کامل [PDF 946 kb]
(745 دریافت)
| چکیده (HTML) (2164 مشاهده)
متن کامل: (1172 مشاهده)
مقدمه
با افزایش جمعیت سالمندان، بروز بیماریهای مزمن مانند دیابت، سالانه افزایش مییابد که یک چالش بزرگ برای سیستم مراقبتهای بهداشت عمومی است (۱). نفروپاتی دیابتی یک عارضه شایع در بیماران مبتلا به دیابت است و یکی از علل اصلی بیماری مزمن کلیوی (Chronic Kidney Disease, CKD) و مرحله نهایی بیماری کلیوی (End-Stage Renal Disease, ESRD) یا نارسایی کلیوی است (۲). تقریباً 40 درصد از بیماران دیابتی، به نفروپاتی دیابتی مبتلا میشوند که بخش قابلتوجهی از آنها به مرحله نهایی بیماری کلیوی میرسند که با افزایش خطر بیماریهای قلبی عروقی و کاهش قابل توجه امید به زندگی همراه است (۳،۴). فاکتور رشد تغییردهنده بتا ۱ (TGF-β1) یک عضو پلیپپتیدی از ابرخانواده سایتوکاینهای TGF-β است. در بیماران دیابت نوع یک و نوع دو، سطح پلاسمایی TGF-β1 بهطور قابلتوجهی افزایش مییابد و در افراد مبتلا به نفروپاتی دیابتی این افزایش بیشتر است (5،6). TGF-β1 تنظیمکننده مهمی برای فیبروز مرتبط با نفروپاتی دیابتی است که با افزایش بیان TGF-β1 کلیه در شرایط هایپرگلیسمی دائمی بیماران دیابتی و مدلهای حیوانی همراه است. همچنین مشاهده شده که در ادرار بیماران مبتلا به نفروپاتی دیابتی سطح بالایی ازTGF-β وجود دارد که نشان دهنده دخالت TGF-β کلیه در پاتوژنز نفروپاتی دیابتی است (۷). تجزیه و تحلیل همبستگی نشان میدهد که سطوح پلاسمایی TGF-β1 با شدت اختلال عملکرد کلیه در بیماران نفروپاتی دیابتی ارتباط نزدیکی دارد (۵). در مجموع این یافتههای بالینی، پاتولوژیک و اپیدمیولوژیک از بیماران دیابتی و مدلهای حیوانی نشان میدهند که TGF-β1 در بیماریزایی نفروپاتی دیابتی نقش دارد. در سطح سلولی نشان داده شده که سیگنالینگ TGF-β نقش مهمی هم در پیری سلولی و پیری سلولهای بنیادی دارد (۸). به طوریکه مسدودکردن TGF-β با آنتیبادی خنثیکننده، اولیگونوکلئوتیدهای ضد حسگر، مهارکنندهها یا حذف ژنتیکی گیرندهها میتواند باعث کاهش فیبروز کلیه شود (۹). از طرف دیگر، Smad7 (Mothersagainst decapentaplegic homolog 7) به عنوان یک تنظیمکننده مهاری در مسیر سیگنالینگ TGF-β/ SMAD شناخته شده است. SMAD7 میتواند توسط یک مکانیسم وابسته به Smad3 القا شود، که به نوبه خود انتقال سیگنال TGF-β1 را از طریق حلقه بازخورد منفی آن مسدود میکند (۱۰). مطالعات اندکی اهمیت ورزش بر فیبروز ناشی از نفروپاتی دیابتی را گزارش کردهاند (۱4-۱1). مطالعات نشان دادند که فعالیت ورزشی میتواند از پیشرفت نفروپاتی دیابتی در موشهای صحرایی جلوگیری کند (۱3-۱1). همچنین فعالیت ورزشی با شدت متوسط با تقویت Sirt1 (Sirtuin 1) و تنظیم EMT (EPithelial-Mesenchymal Transition) ناشی از TGF-β در دیابت نوع دو از پیشرفت بیماری کلیوی جلوگیری میکند (۱۴). Sirt1 نقش مهمی در هموستاز (Homeostasis) گلوکز و حساسیت به انسولین در کبد، عضلات و بافت چربی دارد (۱۵). بیان این پروتئین با فعالیت آنتی دیابتیک، تنظیم ترشح انسولین، بهبود مسیر سیگنالینگ انسولین و عملکرد میتوکندری، مقاومت به استرس و التهاب ارتباط دارد (۱۵). تمرین با بار فزاینده بر فیبروز کلیوی ناشی از پیری طبیعی، با تنظیم مسیر سیگنالینگ غیر SMAD توسط TGF-β و تحریک فعالشدن اتوفاژی در موشهای مسن فیبروز کلیوی را بهبود میبخشد (۱۶). تمرین تناوبی شدید (High-Intensity Interval Traning, HIIT) ترکیبی از تکرار دورههای فعالیت با شدت بالا (نزدیک به بیشینه یا فوق بیشینه) است که با فعالیت کم شدت تا متوسط و در برخی موارد غیر فعال بودن همراه است (۱۷). HIIT در مقایسه با تمرینات با شدت متوسط علیرغم حجم تمرین کمتر منجر به سازگاریهای مشابه یا حتی بیشتر میشود (۱۸). با وجود مطالعات محدود در رابطه با تاثیر فعالیت ورزشی بر فیبروز کلیوی سالمندان دیابتی و با توجه به اینکه TGF-β هم در سالمندی و هم در بیماری دیابت نقش دارد، بنابراین با مهار مسیرTGF-β1/ SMAD ممکن است فیبروز کلیوی دیابت درمان شود (۱۹). با توجه به اینکه فعالیت ورزشی به عنوان یک راهبرد غیر دارویی در درمان دیابت میباشد، پژوهش حاضر با هدف بررسی تاثیر شش هفته HIIT بر تغییرات بیان ژنهای TGF‐β1 و SMAD7 در موشهای صحرایی دیابتی سالمند مورد ارزیابی قرار گرفت.
روش بررسی
در این پژوهش ۳۶ سر موش صحرایی نر نژاد ویستار به عنوان نمونه پژوهش خریداری شدند. سن موش¬های صحرایی ۲۱ ماه و وزن آنها ۳۴۰ تا ۳۹۰ گرم بود. موشها به روش تصادفی ساده به۳ گروه: کنترل سالم (n=12)، کنترل دیابت (n=12) ، تمرین- دیابت (n=12)، تقسیم شدند. بعد از تایید دیابتی شدن، موشهای صحرایی در قفسهای پلیکربنات مجزا در دمای 22±۲ درجه سانتیگراد و چرخه روشنایی-تاریکی ۱۲ :۱۲ ساعت قرار گرفتند. آشناسازی با پروتکل تمرین تناوبی شدید به مدت یک هفته صورت گرفت. تمام حیوانات دسترسی آزاد به آب و غذا داشتند. القا دیابت موشهای صحرایی: در این پژوهش جهت القا دیابت نوع دو در موشها، محلول استرپتوزوتوسین (STZ) (حلشده در بافر سیترات 0/1 مولار با PH=4/5) به صورت داخل صفاقی فقط یک مرتبه با دوز ۶۰ میلیگرم به ازای هر کیلوگرم از وزن بدن بعد از ۱۵ دقیقه تزریق نیکوتینآمید با دوز ۱۱۰ میلیگرم به ازای هر کیلوگرم از وزن بدن تزریق شد. ۷۲ ساعت بعد از تزریق، قند خون موشها با خونگیری از طریق بریدن نوک دم، بهوسیله گلوکومتر اندازهگیری شد و قند خون ناشتای بالای ۲۰۰ میلیگرم بر دسیلیتر به عنوان دیابت نوع ۲ در نظر گرفته شد. به منظور اطمینان از دیابتی شدن موشهای صحرایی یک هفته بعد قند خون آنها مورد بررسی قرار گرفت و موشهای صحرایی با قند خون ناشتای بالای ۲۰۰ میلیگرم بر دسیلیتر به عنوان مدل دیابت نفروپاتی در نظر گرفته شدند (۲۰،۲۱). آزمون تعیین حداکثر اکسیژن مصرفی: جهت تعیین حداکثر اکسیژن مصرفی از آزمون فزاینده استاندارد Bedford و همکاران (۱۹۷۹) استفاده شد که توسط Leandro و همکاران (۲۰۰۷) برای موشهای صحرایی نژاد ویستار استانداردسازی شده است (۲۲،۲۳). آزمون فزاینده به این صورت بود که موشهای صحرایی بر روی تردمیل با سرعت پنج (متر بر دقیقه) شروع به دویدن کردند و هر سه دقیقه سرعت تردمیل (پنج متر بر دقیقه) افزایش یافت. آزمون تا لحظه رسیدن موشهای صحرایی به واماندگی ادامه داشت. سرعت نهایی موشهای صحرایی به عنوان سرعت بیشینه در زمان رسیدن به حداکثر اکسیژن مصرفی برای محاسبه شدتهای تمرینی موشهای صحرایی استفاده شد.
پروتکل تمرین تناوبی شدید: پروتکل تمرین تناوبی شدید بر مبنای پروتکل تمرین تناوبی شدید رضایی و همکاران طراحی شد با این حال با توجه به تفاوت سنی و دیابتی بودن حیوانات و انجام پایلوت اولیه، پروتکل با توجه به توانایی حیوانات تعدیل شد (۲۴). پروتکل تمرین تناوبی شدید شامل سه قسمت گرمکردن، تمرین (شامل تکرار تناوبها) و سردکردن بود. گرمکردن و سرد کردن موشهای صحرایی با شدت ۴۰ درصد VO2max به مدت ۵ دقیقه دویدن بر روی نوارگردان انجام شد. تمرین تناوبی شامل ترکیبی از تکرار¬های تناوب با شدت بالا و پایین بود. تکرار تناوب با شدت بالا شامل ۲ دقیقه دویدن با شدت ۶۰ درصد VO2max در هفته اول، ۷۰ درصد VO2max در هفته دوم، ۸۰ درصد VO2max در هفته سوم و ۹۰ درصد VO2max از ابتدای هفته چهارم تا پایان هفته ششم بود. تکرار تناوب با شدت پایین در تمام هفتهها شامل ۲ دقیقه دویدن با شدت ۴۰ درصد VO2max بود. تمرین تناوبی به گونه¬ای بود که پس از گرم کردن، موشهای صحرایی ابتدا تناوب با شدت بالا و سپس تناوب با شدت پایین را اجرا کردند. پس از انجام آخرین تکرار تناوب با شدت بالا، موشهای صحرایی به مدت ۵ دقیقه دویدن با شدت40 درصد VO2max سردکردن را انجام دادند. تعداد تکرار تناوب با شدت بالا با توجه به هفته تمرینی موشهای صحرایی تعیین شد. به طوریکه در هفته اول 3 تکرار تناوب، هفته دوم ۵ تکرار تناوب، هفته سوم ۷ تکرار تناوب و از ابتدای هفته چهارم تا پایان هفته ششم شامل ۹ تکرار تناوب بود. از این رو زمان کل تمرین شامل تکرار تناوب با شدت بالا، تکرار تناوب با شدت پایین به همراه گرمکردن و سردکردن در هفته اول ۲۰ دقیقه، در هفته دوم ۲۸ دقیقه، هفته سوم ۳۶ دقیقه و از ابتدای هفته چهارم به بعد ۴۴ دقیقه بود. شیب دستگاه در کل پروتکل تمرینی صفر در نظر گرفته شد. (مطابق با جدول ۱). گروههای کنترل سالم و دیابت در این مدت هیچ گونه تمرینی انجام ندادند (24-22).
اندازهگیری میزان بیان ژنهای TGF‐β1 و SMAD7: بیست و چهار ساعت پس از آخرین جلسه تمرینی، موشها با ترکیبی از کتامین (mg/kg۹۰) و زایلازین (mg/kg۱۰) بیهوش شدند (۲۵). بافت کلیه تحت شرایط استریل جدا شد و بلافاصله در نیتروژن مایع (دمای ۱۹۶-) منجمد شد و تا زمان فرا رسیدن انجام کار استخراج RNA در فریزر ۸۰- درجه نگهداری شدند. برای بررسی بیان ژنهای TGF-1 و SMAD7 در بافت کلیه موشهای صحرایی با استفاده از کیت ستونی استخراج RNA (FavorPreP™ Tissue Total RNA Mini Kit) ساخت کشور هنگکنگ طبق دستورالعمل (FATRK 001)، کل محتویات RNA سلول (total RNA) استخراج شد. بافت با استفاده از یک میلیمول RNAx هموژن شد سپس با اضافه کردن ترکیبی از RB بافر و بتامرکاپتواتانول (MercaPtoethanol -) نمونه لیز شده و با دستگاه همگن کننده بافت هموژن گردید. در مرحله بعد نمونه روی فیلتر سفید گذاشته شد و به مدت ۳۰ دقیقه سانتریفیوژ گردید. ماده زیر توری سفید بدون رسوب ایجاد شده در آن به میکروتیوب دیگری منتقل شد. RNA استخراج شده با ۴۵۰ میکرولیتر اتانول ۷۰% شستشو و خشک گردید و سپس 1/500 میکرولیتر آب اضافه شد و ۳۰ دقیقه سانتریفیوژ گردید. در مرحله بعد ۷۵۰ میکرولیتر آب استریل اضافه و ۳۰ دقیقه سانتریفیوژ گردید. این مرحله دو مرتبه تکرار شد. سپس RNAase free water اضافه شد و ۳۰ دقیقه با دور بالا سانتریفیوژ شد. کمیت و کیفیت RNA استخراج شده با استفاده از اسپکتروفتومتری نانو دراپ با نسبت چگالی نوری 280/260 نانومتر و الکتروفورز بر روی ژل آگارز (1%) ارزیابی شد (دستگاه پیکو دراپ شرکت سیگما ساخت امریکا). سنتز cDNA طبق دستورالعمل موجود در کیت فرمنتاز (K1621) تهیه گردید. هنگام تهیه cDNAاز نمونه تخلیص شده پس از قرائت جذب، حجمی شامل ۱۰۰۰ نانوگرم RNA برداشته شد، سپس 0/5 میکرولیتر رندوم هگزامر (Random Hexamers)، 0/5 میکرولیتر پرایمر الیگو دی تی (Oligo dT) به آن افزوده و تا حجم ۱۲ میکرولیتر آب DEPC اضافه گردید و به دمای 65 درجه به مدت 5 دقیقه منتقل شد. سپس به مدت 2 دقیقه بر روی یخ قرار گرفت تا ساختارهای ثانویه RNA از هم باز شوند. در مرحله بعد ۴ میکرولیتر 5X Reaction Buffer، ۲ میکرولیتر dNTP، ۱ میکرولیتر RiboLock RNase Inhibitorو ۱ میکرولیتر RevertAid RT به ترکیب قبل اضافه گردید. سپس ترکیب ابتدا به مدت 5 دقیقه در دمای 25 درجه قرار گرفت. بعد از آن به مدت 60 دقیقه در دمای 42 درجه قرار گرفت. در آخر به منظور از کار افتادن آنزیم RT، تیوبهای واکنش به مدت 5 دقیقه در دمای 70 درجه سانتیگراد قرار گرفت. سپس cDNA آماده شده جهت انجام RT- PCR مورد استفاده قرار گرفت. تمام پرایمرها توسط نرمفزار Allele IDv7.8 طراحی شد و از ژن خانهبان (HousekeePing) B2m به عنوان کنترل داخلی واکنشهای qPCR استفاده شد. تمام پرایمرها به صورت اتصال اگزون- اگزون طراحی شدند. جهت اطمینان از عدم تکثیر DNA ژنومی از 25 نانوگرم cDNA و 25 نانوگرم RNA در تیوبهای جداگانه از واکنش PCR و بهکارگیری از ژل آگاروز 5/1% استفاده شد. همه نمونهها برای شرایط qPCR آماده شدند و در دمای 94 درجه سانتیگراد به مدت 10 دقیقه به دنبال 40 سیکل 15 ثانیهای در دمای 94 درجه سانتیگراد، 60 ثانیه در دمای 58 درجه سانتیگراد و سپس 7 دقیقه در دمای 72 درجه سانتیگراد قرار گرفتند (۲۶). برای کمی سازی مقادیر بیان ژن از فرمول ct-۲ (۲به توان منفی ct) استفاده شد و مقادیر Fold change محاسبه شد (۲۷). تمام آزمایشات RT-PCR در سه تکرار انجام شد. توالی پرایمرهای پژوهش در جدول ۲ ارائه شده است.
تجزیه و تحلیل آماری
برای بررسی طبیعی بودن توزیع دادهها از آزمون شاپیرو-ویلک و جهت بررسی تفاوت معناداری بین گروهها از آزمون تحلیل واریانس آنوا یک طرفه و آزمون تعقیبی توکی استفاده شد. تجزیه و تحلیلهای آماری با استفاده از نرمافزارSPSS version 16 در سطح معناداری P<0/05 انجام شد.
ملاحظات اخلاقی
پروپوزال این پژوهش، مورد تایید کمیته اخلاق در پژوهش دانشگاه علوم پزشکی شیراز (IR.SUMS.REHAB.REC.1400.027) قرار گرفته است.
نتایج
نتایج تحلیل واریانس آنوای یک طرفه نشان داد که بهطور کلی بین وزن بدن موشهای صحرایی در گروههای مختلف تفاوت معناداری وجود دارد (P= 0/000، F=26/47). القای دیابت منجر به کاهش وزن بدن موشهای صحرایی شد (P=0/000)، همچنین بین وزن موشهای صحرایی در گروههای دیابت تمرین و کنترل دیابت تفاوت معناداری وجود نداشت (P=0/13). نتایج آزمون تعقیبی توکی در جدول ۳ نشان داده شده است. نتایج تحلیل واریانس آنوای یک طرفه تفاوت معناداری را در بیان ژن TGF-β1 بافت کلیه در بین گروه¬های مورد مطالعه نشان داد (P=0/000، F= 34/39) (جدول 4). نتایج آزمون تعقیبی توکی نشان داد که گروه تمرین- دیابت کاهش معناداری در بیان ژن TGF-β1 بافت کلیه نسبت به گروه دیابت داشت (P=0/001). همچنین القا دیابت منجر به افزایش بیان ژن TGF-β1 (P=0/000) در بافت کلیه موش¬های صحرایی سالمند نسبت به گروه کنترل سالم شد (0/001=P) (شکل ۱). علاوه بر این تفاوت معناداری در بیان ژن SMAD7 در بافت کلیه در بین گروههای مورد مطالعه وجود داشت (P=0/000، F= 17/84) (جدول 5). نتایج آزمون تعقیبی توکی نشان داد که گروه تمرین- دیابت نسبت به گروه کنترل- دیابت منجر به افزایش معناداری در بیان ژن SMAD7 در بافت کلیه شد (0/008=P). همچنین بین گروه دیابت و کنترل سالم در بیان ژن SMAD7 کاهش معناداری مشاهده شد (0/001=P) (شکل۲).
جدول ۱: پروتکل تمرینی مورد استفاده در پژوهش

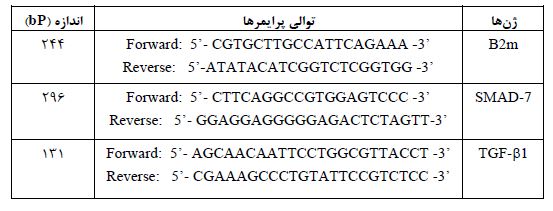
جدول ۲: توالی پرایمرهای استفاده شده در این پژوهش
جدول ۳: میانگین ± انحراف معیار وزن بدن موشهای صحرایی (گرم) و نتایج آزمون تعقیبی توکی در گروههای مختلف

علامت * نشان دهنده تفاوت معنادار است
جدول4: نتایج تحلیل واریانس آنوای یک طرفه TGF-1
.JPG)
علامت * نشان دهنده تفاوت معنادار است
.JPG)
شکل۱: تغییرات بیان mRNA TGF-β1 در گروه¬های کنترل سالم (Control)، کنترل دیابت (Diabetes) و دیابت- تمرین (Diabetes+ HIIT).
علامت * نشان دهنده تفاوت معنادار است
جدول5: نتایج تحلیل واریانس آنوای یک طرفه SMAD7

(Smad7 (Mothers against decapentaplegic homolog 7 علامت * نشان دهنده تفاوت معنادار است
.JPG)
شکل۲: تغییرات بیان mRNA SMAD7 در گروههای کنترل سالم (Control)، کنترل- دیابت (Diabetes) و دیابت- تمرین (Diabetes+ HIIT). علامت * نشان دهنده تفاوت معنادار است.
بحث
با تغییر سبک زندگی و روند پیری جمعیت، بروز دیابت نوع دو سال به سال در حال افزایش است. شیوع دیابت یک مشکل رو به رشد بهداشت عمومی جهانی است (۲۸). نفروپاتی دیابتی یک عارضه میکروواسکولار (Microvascular) جدی در بیماران دیابتی است که با گلومرولواسکلروزیس (Glomerulosclerosis) ناشی از متابولیسم غیرطبیعی گلوکز مشخص میشود و همچنین علت اصلی مرگ در نفروپاتی دیابتی است (۲۹). پیشرفت این بیماری به تدریج میتواند منجر به کاهش عملکرد کلیوی، ادم سیستمیک و حتی نارسایی کلیه در مرحله پایانی تبدیل شود که یکی از علل شایع مرگ در بیماران دیابتی است (۳۰،۳۱). گرچه ورزش آثار مفید مختلفی در دیابت نوع دو و چاقی دارد، اما اثرات کلیوی ورزش، بهخصوص HIIT و مکانیسمهای زیربنایی مسئول هنوز به طور کامل مشخص نشده است. پژوهش حاضر به منظور بررسی تاثیر شش هفته تمرین تناوبی شدید بر بیان ژن TGF-β1 و SMAD7 در بافت کلیه در موشهای صحرایی سالمند دیابتی صورت گرفت. نتایج مطالعه نشان داد که تمرین تناوبی شدید آثار محافظتی در برابر نفروپاتی دارد. در پژوهش حاضر برای شناخت تغییرات فیزیولوژیکی بافت کلیه در پاسخ به تمرینات تناوبی شدید، TGF-β1 مورد بررسی قرار گرفت. نتایج پژوهش نشان داد که HIIT منجر به کاهش معناداری در بیان ژن TGF-β1 در بافت کلیه میشود. همسو با این نتایج Ren و همکاران (۲۰۱۷) نشان دادند که فعالیت ورزشی با شدت متوسط با تقویت Sirt1 و تنظیم EMT ناشی از TGF-β در دیابت نوع دو از پیشرفت بیماری کلیوی جلوگیری میکند (۱۴). به علاوه BAO و همکاران (۲۰۱۹) تاثیر تمرین با بار فزاینده بر فیبروز کلیوی ناشی از پیری طبیعی و مکانیسمهای آن را مورد مطالعه قرار دادند و نشان دادند که تمرین فزاینده با تنظیم مسیر سیگنالینگTGF‐β1/ TAK1/ MKK3/ P38MAPK (مسیرسیگنالینگ غیر SMAD) و تحریک فعالشدن اتوفاژی در موشهای مسن فیبروز کلیوی را بهبود میبخشد (۱۶). با وجود این برخی تحقیقات افزایش سطح TGF-β را بر اثر تمرین در بافتهای دیگر گزارش کردهاند (۳۲،۳۳). مقدم و همکاران (۱۳۹۶) اثر حفاظتی تمرین هوازی بر سرطان پستان بهواسطه پروتئین TGF-β در موشهای ماده را مورد ارزیابی قرار دادند. نتایج آنها نشان داد که ده هفته تمرین استقامتی هوازی منجر به افزایش معنادار میانگین مقادیر پروتئین TGF-β در بافت تومور نسبت به گروه کنترل شد (۳۳). احتمالاً دلیل افزایش TGF-β متفاوت بودن نوع تمرین باشد. در پژوهش دیگری نشان داده شد که هشت هفته تمرین هوازی منجر به افزایش مقدار TGF-β در بافت قلب موشهای صحرایی سالمند میشود (۳۲). این افزایش شاید به دلیل کافی نبودن شدت در برنامه تمرینی مطالعه فوق باشد. نشان داده شده که هایپرگلیسمی حاد و مزمن بیان TGF-β1 را القا میکند. TGF-β1 یک سایتوکاین اصلی در چاقی و مقاومت به انسولین است و همچنین با داشتن خواص فیبروژنیک نقش محوری در ایجاد نفروپاتی دیابتی دارد (۳۴). عوامل موثر بر بیان TGF-β1 بیشمار هستند (۳۵). هایپرگلیسمی یکی از عوامل اصلی بیان TGF-β1 است و بیماران دیابتی سطح TGF-β1 بالاتری نسبت به افراد سالم دارند. تصور میشود که مسیر دی آسیل گلیسرول پروتئین کیناز C، مسیر گلوکوزآمین و احتمالاً مسیر پلیول، نقش مهمی در تولید بیشتر TGF-β1در دیابت دارند (۳۶). یافتههای محققان حاکی از این است که گلوکز واسطه اصلی بیان TGF-β1است (۳۷). هایپرگلیسمی که معمولاً در بیماران مبتلا به دیابت نوع دو مشاهده میشود با بیش تنظیمی انتقالدهنده گلوکز-۱ (GLUT1) همراه است که منجر به بیان بیش از حد TGF-β توسط سلولهای توبولی مزانژیال یا سلولهای کلیوی میشود (۳۸،۳۹). علاوه بر این افزایش فشار داخل گلومرولی، کشش سلول مزانژیال، فعال شدن سیستم رنین-آنژیوتانسین، گونههای فعال اکسیژن (ROS) و محصولات نهایی گلیکاسیون پیشرفته (AGEs) احتمالاً باعث تولید TGF-β در کلیه یا در سلولهای مزانژیال یا توبولی میشود (۳۹،۴۰). TGF-β از طریق مسیرهای اتوکرین و پاراکرین عمل کرده و گیرندههای سیگنالینگ و سنتز ECM آن را تحریک میکنند (۳۹،۴۰). شواهد بالینی نشان میدهد که پاسخ ناشی از TGF-β شامل یک مسیر نهایی است که منجر به توسعه گلومرولو اسکلروز و فیبروز بینابینی در مرحله پایانی بیماری کلیوی (ESRD) میشود (۴۱). با توجه به مکانسیم فوق کاهش میزان هایپرگلیسمی و مقاومت به انسولین در گروه HIIT شاید منجر به کاهش بیان TGF-β1 در بافت کلیه شود که بیانگر این است که تمرین تناوبی شدید میتواند اثرات مخرب مسیر سیگنالینگ TGF-β1 را کاهش داده و از شدت آسیب به بافت کلیه در بیماران نفروپاتی دیابتی جلوگیری کند. یکی دیگر از نتایج پژوهش حاضر افزایش معنادار بیان ژن SMAD7 در بافت کلیه در پاسخ به HIIT بود. مطالعات اندکی تاثیر تمرینات ورزشی را بر بیان mRNA SMAD7 در بافت کلیه در بیماری دیابت را بررسی کردهاند. در این راستا Hou و همکاران (۲۰۲۱) نشان دادند که تمرینات شنا به مدت ۱۲ هفته بیان mRNA SMAD7 میوکارد در موش-های صحرایی دیابتی را افزایش میدهد و منجر به کاهش معناداری در قند خون و بیان mRNA TGF-β1 میشود (۴۲). همچنین Wang و Yuan (۲۰۱۹) گزارش کردند که مدل تمرین تناوبی سرعتی (SIT) میتواند منجر به تنظیم مثبت SMAD7 در بافت کلیه موشهای صحرایی دیابتی شود (۴۳). اجزای اساسی سیگنالینگ TGF-β1 شامل یک کمپلکس گیرنده با گیرندههای مرتبط با غشا نوع I و نوع II و پروتئینهای SMAD است که در عوامل رونویسی پایین دستی نقش دارند (۴۴،۴۵). SMAD7به عنوان یک تنظیم کننده مهاری در مسیر سیگنالینگ TGF-β/ SMAD میتواند توسط یک مکانیسم وابسته به Smad3 القا شود، که به نوبه خود انتقال سیگنال TGF-β1 را از طریق حلقه بازخورد منفی آن مسدود میکند. SMAD7 با مسدود کردن دسترسی R-Smadها به TβRI و یا با افزایش تخریب کمپلکسهای گیرنده، فسفوریلاسیون R-Smad را مهار میکنند (۱۰). در مقابل شکل فعال TGF-β، شکل نهفته TGF-β1 میتواند با بیش تنظیم SMAD7 در موشهای ترانسژنیک از کلیهها در برابر فیبروز و التهاب محافظت کند (۱۰). گزارشات نشان میدهد که القای SMAD7 گذار اپیتلیال مزانشیمی توبولی و ایجاد آسیبهای فیبروتیک را بلاک میکند (۴۶). نقش عملکردی SMAD7 بیشتر با یافتههایی مشخص میشود که حذف SMAD7 باعث تسریع فیبروژنز کلیه در نفروپاتی انسدادی، نفروپاتی دیابتی و همچنین نفروپاتی فشار خون میشود (۴۹-۴۷)، که SMAD7 را به عنوان یک عامل درمانی برای درمان بیماریهای مزمن کلیوی نشان میدهد (49-52،۴7). SMAD7 سیگنالهای TGF-β/ SMAD را از طریق دو مکانیسم ممکن بهطور منفی تنظیم میکند. نخست اینکه SMAD7 به گیرنده TGF-β1 متصل میشود و در نتیجه از جذب و فسفوریلاسیون Smad2 و Smad3 جلوگیری میکند و دیگر اینکه SMAD7به عنوان یک گیرنده عمل میکند (۵۳). پروتئین آداپتوری که لیگازهای یوبیکوئیتین مانند Smurf2 را به مجموعه گیرنده TGF-β جذب میکند تا تخریب آن را از طریق مسیرهای تخریب پروتئازوم- یوبیکوئیتین تقویت کند (۵6-۵4). با توجه به مکانسیم فوق، تمرین تناوبی شدید با کاهش بیان TGF-β1 در بافت کلیه و تاثیرش بر افزایش بیان SMAD7 میتواند منجر به اتصال SMAD7 به گیرنده TGF-β1 شده و فسفریلاسیون R-Smad را مهار کند و یا با افزایش تخریب کمپلکسهای گیرنده از آسیبهای فیبروتیک جلوگیری کند. بنابراین با توجه به نتایج پژوهش حاضر شاید HIIT بهتواند به عنوان یک روش درمانی غیر دارویی در جلوگیری از پیشرفت نفروپاتی دیابتی، از طریق مسدود کردن فیبروز کلیه با واسطه TGF-β1 و تنظیم مثبت SMAD7 عمل کند که ممکن است اساس درمان بیماران نفروپاتی دیابتی را در آینده فراهم کند. از آنجایی که AGEs یک واسطه ضروری در عوارض دیابت هستند، این محصولات قادر به فعالکردن Smad 2/3 به طور مستقیم و مستقل از TGF-β هستند و توسط گیرنده AGEs از طریق مسیر تداخلی وابسته به MAP کیناز ERK/P38 عمل میکنند. با توجه به اینکه مطالعه حاضر تاثیر HIIT بر مسیر TGF-β /Smad را مورد بررسی قرار داده لذا لازم به ذکر است که در پژوهشهای آینده نیاز است تا تاثیر HIIT بر مسیر ERK/P38 در موشهای دیابتی سالمند مورد بررسی قرار گیرد. از محدودیتهای این پژوهش میتوان به عدم اندازهگیری گیرنده TGF-β1 و فاکتورهای درگیر در مسیر غیروابسته به SMAD اشاره کرد.
نتیجهگیری
بهطور کلی مطالعه حاضر نشان داد که القای دیابت منجر به افزایش معناداری در بیان mRNA TGF-β1 و کاهش معناداری در بیان mRNA SMAD7 میشود و شش هفته تمرین تناوبی شدید به ترتیب منجر به کاهش و افزایش معنادار در mRNA TGF-β1 و mRNA SMAD7 در بافت کلیه موش¬های صحرایی دیابتی میشود و در نتیجه میتواند در کلیهها اثرات محافظتی در برابر نفروپاتی را اعمال کند.
سپاسگزاری
مقاله حاضر برگرفته از رساله دکتری دانشجویی دانشگاه شیراز میباشد، لذا نویسندگان مراتب تقدیر و تشکر خود را از کلیه افرادی که در اجرای این پژوهش ایفای نقش کردند، اعلام میدارند.
حامی مالی: ندارد.
تعارض در منافع: وجود ندارد.
References:
1- Shlisky J, Bloom DE, Beaudreault AR, Tucker KL, Keller HH, Freund-Levi Y, et al. Nutritional Considerations for Healthy Aging and Reduction in Age-Related Chronic Disease. Adv in Nutr 2017; 8(1): 17-26.
2- Chang AS, Hathaway CK, Smithies O, Kakoki M. Transforming Growth Factor-Β1 and Diabetic Nephropathy. American J Physiology-Renal Physiology 2016; 310(8): 689-96.
3- Association AD. Standards of Medical Care in Diabetes-2014. Diabetes Care 2014; 37: 14-80.
4- Dekkers CC, Gansevoort RT, HeersPink HJ. New Diabetes Therapies and Diabetic Kidney Disease Progression: The Role of SGLT-2 Inhibitors. Current Diabetes Reports 2018; 18(5): 1-12.
5- Ibrahim S, Rashed L. Estimation of Transforming Growth Factor-Beta 1 as a Marker of Renal Injury in Type II Diabetes Mellitus. Saudi Med J 2007; 28(4): 519-23.
6- Jakuš V, SaPák M, Kostolanská J. Circulating TGF-Β1, Glycation, and Oxidation in Children with Diabetes Mellitus Type 1. ExP Diabetes Res 2012; 2012: 510902.
7- Shaker YM, Soliman HA, Ezzat E, Hussein NS, Ashour E, Donia A, et al. Serum and Urinary Transforming Growth Factor Beta 1 as Biochemical Markers in Diabetic Nephropathy Patients. Beni-Suef University J Basic and APPlied Sciences 2014; 3(1): 16-23.
8- Tominaga K, Suzuki HI. TGF-Β Signaling in Cellular Senescence and Aging-Related Pathology. International J Molecular Sciences 2019; 20(20): 5002.
9- Hills CE, Squires PE. The Role of TGF-Β and Epithelial-To Mesenchymal Transition in Diabetic Nephropathy. Cytokine Growth Factor Rev 2011; 22(3): 131-9.
10- Meng X-M, Tang PM-K, Li J, Lan HY. TGF-Β/Smad Signaling in Renal Fibrosis. Front Physiol 2015; 6: 82.
11- Boor P, Celec P, Behuliak M, Grančič P, Kebis A, Kukan M, et al. Regular Moderate Exercise Reduces Advanced Glycation and Ameliorates Early Diabetic Nephropathy in Obese Zucker Rats. Metabolism 2009; 58(11): 1669-77.
12- Chiasera JM, Ward-Cook KM, McCune SA, Wardlaw GM. Effect of Aerobic Training on Diabetic Nephropathy in a Rat Model of Type 2 Diabetes Mellitus. Ann Clin Lab Sci 2000; 30(4): 346-53.
13- Ito D, Cao P, Kakihana T, Sato E, Suda C, Muroya Y, et al. Chronic Running Exercise Alleviates Early Progression of Nephropathy with Upregulation of Nitric Oxide Synthases and Suppression of Glycation in Zucker Diabetic Rats. PloS one 2015; 10(9): e0138037.
14- Ren L, Sen U, PushPakumar S. Exercise Training Reduces TGF‐Β Mediated Epithelial Mesenchymal Transition in Diabetic Kidney. The FASEB Journal 2017; 31(S1): 1086-5.
15- Cantó C, Auwerx J. PGC-1alpha, SIRT1 and AMPK, an Energy Sensing Network that Controls Energy Expenditure. Curr Opin Lipidol 2009; 20(2): 98-105.
16- Bao C, Yang Z, Cai Q, Li Q, Li H, Shu B. Incremental Load Training Improves Renal Fibrosis by Regulating the TGF‑Β1/TAK1/MKK3/P38MAPK Signaling Pathway and Inducing the Activation of Autophagy in Aged Mice. International J Molecular Med 2019; 44(5): 1677-86.
17- Laursen PB, Jenkins DG. The Scientific Basis for High-Intensity Interval Training. Sports Med 2002; 32(1): 53-73.
18- Winding KM, Munch GW, IePsen UW, Van Hall G, Pedersen BK, Mortensen SP. The Effect on Glycaemic Control of Low Volume High Intensity Interval Training Versus Endurance Training in Individuals with Type 2 Diabetes. Diabetes, Obesity and Metabolism 2018; 20(5): 1131-9.
19- Wu W, Huang XR, You Y, Xue L, Wang X-J, Meng X, et al. Latent TGF-Β1 Protects Against Diabetic Kidney Disease Via Arkadia/Smad7 Signaling. Int J Biol Sci 2021; 17(13): 3583-94.
20- Pierre W, Gildas AJH, Ulrich MC, Modeste WN, Albert K. Hypoglycemic and Hypolipidemic Effects of Bersama Engleriana Leaves in Nicotinamide/Streptozotocin-Induced Type 2 Diabetic Rats. BMC Complement Altern Med 2012; 12(1): 264.
21- Maheshwari R, Balaraman R, Sen AK, Shukla D, Seth A. Effect of Concomitant Administration of Coenzyme Q10 with Sitagliptin on Experimentally Induced Diabetic Nephropathy in Rats. Ren fail 2017; 39(1): 130-9.
22- Bedford TG, TiPton CM, Wilson NC, OPPliger RA, Gisolfi CV. Maximum Oxygen Consumption of Rats and Its Changes with Various Experimental Procedures. J Appl Physiol Respir Environ Exerc Physiol 1979; 47(6):1278-83.
23- Leandro C G, Levada AC, Hirabara SM, Manhães-de-Castro R, De-Castro CB, Curi R, et al. A Program of Moderate Physical Training for Wistar Rats Based on Maximal Oxygen Consumption. J Strength and Cond Res 2007; 21(3): 751-6.
24- Rezaei R, Nasoohi S, HaghParast A, Khodagholi F, Bigdeli MR, Nourshahi M. High Intensity Exercise Preconditioning Provides Differential Protection Against Brain Injury Following Experimental Stroke. Life Sciences 2018; 207: 30-5.
25- Kumar AH, Clover AJ. Intraperitoneal Co-Administration of Low Dose Urethane with Xylazine and Ketamine for Extended Duration of Surgical Anesthesia in Rats. Lab Anim Res 2015; 31(4): 174-9.
26- Vafaei H, Kavari G, Izadi HR, Dorahi ZZ, DianatPour M, DaneshParvar A, et al. Wi-Fi (2.4 Ghz) Affects Antioxidant Capacity, DNA Repair Genes Expression and, Apoptosis in Pregnant Mouse Placenta. Iran J Basic Med Sci 2020; 23(6): 833-40.
27- Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real Time Quantitative PCR and the 2− ΔΔCT Method. Methods 2001; 25(4): 402-8.
28- Sugiyama T, Goryoda S, Inoue K, Sugiyama-Ihana N, Nishi N. Construction of a Simulation Model and Evaluation of the Effect of Potential Interventions on the Incidence of Diabetes and Initiation of Dialysis Due to Diabetic Nephropathy in Japan. BMC Health Serv Res 2017; 17(1): 833.
29- Sharma D, Bhattacharya P, Kalia K, Tiwari V. Diabetic Nephropathy: New Insights into Established Therapeutic Paradigms and Novel Molecular Targets. Diabetes Res Clin Pract 2017; 128: 91-108.
30- Warren AM, Knudsen ST, CooPer ME. Diabetic Nephropathy: An Insight Into Molecular Mechanisms And Emerging Therapies. Expert Opin Ther Targets 2019; 23(7): 579-91.
31- Zürbig P, Mischak H, Menne J, Haller H. CKD273 Enables Efficient Prediction of Diabetic Nephropathy in Nonalbuminuric Patients. Diabetes Care 2019; 42(1): e4-e5.
32- Goodarzi F, Abednatanzi H, Nikbakht H, Ebrahim Kh, Ghazaliyan F. Effects of Eight Weeks Aerobic Exercise on the Signaling Pathway of Cardiac Fibrosis in Elderly Rats. Journal of Knowledge & Health in Basic Medical Sciences 2020; 14(4): 48-53.[Persian]
33- Moghadam V, Piri M, Azarbayjani MA, Matinhomaee H. The Protective Effect of Aerobic Exercise on Breast Cancer by Tgfβ Protein and Smad-3 and MMP2 Gene in Female Mice. SJKU 2017; 22(3): 60-73. [Persian]
34- Wang M, Zhang J, SPinetti G, Jiang L-Q, Monticone R, Zhao D, et al. Angiotensin II Activates Matrix Metalloproteinase Type II and Mimics Age-Associated Carotid Arterial Remodeling in Young Rats. Am J Pathol 2005; 167(5):1429-42.
35- Tzavlaki K, Moustakas A. TGF-β Signaling. Biomolecules 2020; 10(3): 487.
36- Rossert J, Terraz-Durasnel C, Brideau G. Growth Factors, Cytokines, and Renal Fibrosis During the Course of Diabetic Nephropathy. Diabetes Metab 2000; 26 Suppl 4: 16-24.
37- HeydarPour F, Sajadimajd S, Mirzarazi E, HaratiPour P, Joshi T, Farzaei MH, et al. Involvement of TGF-Β and Autophagy Pathways in Pathogenesis of Diabetes: A Comprehensive Review on Biological and Pharmacological Insights. Front Pharmacol 2020; 11: 498758.
38- El-Sherbini SM, Shahen SM, Mosaad YM, Abdelgawad MS, Talaat RM. Gene Polymorphism of Transforming Growth Factor-Β1 in Egyptian Patients with Type 2 Diabetes and Diabetic Nephropathy. Acta Biochim BioPhys Sin 2013; 45(4): 330-8.
39- Qian Y, Feldman E, Pennathur S, Kretzler M, Brosius III FC. From Fibrosis to Sclerosis: Mechanisms of Glomerulosclerosis in Diabetic Nephropathy. Diabetes 2008; 57(6): 1439-45.
40- Sharma K, Ziyadeh FN. Hyperglycemia and Diabetic Kidney Disease: The Case for Transforming Growth Factor–Β as a Key Mediator. Diabetes 1995; 44(10):1139-46.
41- Border WA, Noble NA, Yamamoto T, HarPer JR, Yamaguchi Y, Pierschbacher MD, et al. Natural Inhibitor of Transforming Growth Factor-Β Protects Against Scarring in Experimental Kidney Disease. Nature 1992; 360(6402): 361-4.
42- Hou G, Liu Q, Xi X, Liu H. Effects of Swimming And Epigallocatechin Gallate on Interstitial Proteins Expression of Myocardium From Type 2 Diabetic Rats. Wei Sheng yan jiu 2021; 50(1): 86-92.
43- Wang S-Q, Li D, Yuan Y. Long-Term Moderate Intensity Exercise Alleviates Myocardial Fibrosis in Type 2 Diabetic Rats Via Inhibitions of Oxidative Stress and TGF-Β1/Smad Pathway. J Physiol Sci 2019; 69(6): 861-73.
44- Xie F, Ling L, van Dam H, Zhou F, Zhang L. TGF-Β Signaling in Cancer Metastasis. Acta Biochim Biophys Sin (Shanghai) 2018; 50(1):121-32.
45- Zhang YE. Mechanistic Insight Into Contextual TGF-Β Signaling. Curr Opin Cell Biol 2018; 51: 1-7.
46- Saika S, Ikeda K, Yamanaka O, Sato M, Muragaki Y, Ohnishi Y, et al. Transient Adenoviral Gene Transfer of Smad7 Prevents Injury Induced Epithelial Mesenchymal Transition of Lens Epithelium in Mice. Lab invest 2004; 84(10):1259-70.
47- Chen HY, Huang XR, Wang W, Li JH, Heuchel RL, Chung AC, et al. The Protective Role of Smad7 in Diabetic Kidney Disease: Mechanism and Therapeutic Potential. Diabetes 2011; 60(2): 590-601.
48- Chung AC, Huang XR, Zhou L, Heuchel R, Lai KN, Lan HY. Disruption of the Smad7 Gene Promotes Renal Fibrosis and Inflammation in Unilateral Ureteral Obstruction (UUO) in Mice. Nephrol Dial Transplant 2009; 24(5): 1443-54.
49- Liu G-X, Li Y-Q, Huang XR, Wei LH, Zhang Y, Feng M, et al. Smad7 Inhibits Angii-Mediated Hypertensive Nephropathy in a Mouse Model of Hypertension. Clin Sci 2014; 127(3): 195-208.
50- Ka S, Yeh Y, Huang X, Chao T, Hung Y, Yu C, et al. Kidney-Targeting Smad7 Gene Transfer Inhibits Renal TGF-Β/MAD Homologue (SMAD) and Nuclear Factor Κb (NF-Κb) Signalling Pathways, and Improves Diabetic Nephropathy in Mice. Diabetologia 2012; 55(2): 509-19.
51- Ka S-M, Huang X-R, Lan H-Y, Tsai P-Y, Yang S-M, Shui H-A, et al. Smad7 Gene Therapy Ameliorates an Autoimmune Crescentic Glomerulonephritis in Mice. J Am Soc NePhrol 2007; 18(6): 1777-88.
52- Lan HY, Mu W, Tomita N, Huang XR, Li JH, Zhu H-J, et al. Inhibition of Renal Fibrosis by Gene Transfer of Inducible Smad7 Using Ultrasound-Microbubble System in Rat UUO Model. J Am Soc NePhrol 2003; 14(6): 1535-48.
53- Zhu H-J, Iaria J, Sizeland AM. Smad7 Differentially Regulates Transforming Growth Factor Β-Mediated Signaling Pathways. J Biol Chem 1999; 274(45): 32258-64.
54- Ebisawa T, Fukuchi M, Murakami G, Chiba T, Tanaka K, Imamura T, et al. Smurf1 Interacts with Transforming Growth Factor-Β Type I Receptor through Smad7 and Induces Receptor Degradation. J Biol Chem 2001; 276(16): 12477-80.
55- Kavsak P, Rasmussen RK, Causing CG, Bonni S, Zhu H, Thomsen GH, et al. Smad7 Binds to Smurf2 to Form an E3 Ubiquitin Ligase that Targets the Tgfβ Receptor for Degradation. Mol cell 2000; 6(6): 1365-75.
56- Liu F-Y, Li X-Z, Peng Y-M, Liu H, Liu Y-H. Arkadia-Smad7-Mediated Positive Regulation of TGF-Β Signaling in a Rat Model of Tubulointerstitial Fibrosis. Am J Nephrol 2007; 27(2): 176-83.
با افزایش جمعیت سالمندان، بروز بیماریهای مزمن مانند دیابت، سالانه افزایش مییابد که یک چالش بزرگ برای سیستم مراقبتهای بهداشت عمومی است (۱). نفروپاتی دیابتی یک عارضه شایع در بیماران مبتلا به دیابت است و یکی از علل اصلی بیماری مزمن کلیوی (Chronic Kidney Disease, CKD) و مرحله نهایی بیماری کلیوی (End-Stage Renal Disease, ESRD) یا نارسایی کلیوی است (۲). تقریباً 40 درصد از بیماران دیابتی، به نفروپاتی دیابتی مبتلا میشوند که بخش قابلتوجهی از آنها به مرحله نهایی بیماری کلیوی میرسند که با افزایش خطر بیماریهای قلبی عروقی و کاهش قابل توجه امید به زندگی همراه است (۳،۴). فاکتور رشد تغییردهنده بتا ۱ (TGF-β1) یک عضو پلیپپتیدی از ابرخانواده سایتوکاینهای TGF-β است. در بیماران دیابت نوع یک و نوع دو، سطح پلاسمایی TGF-β1 بهطور قابلتوجهی افزایش مییابد و در افراد مبتلا به نفروپاتی دیابتی این افزایش بیشتر است (5،6). TGF-β1 تنظیمکننده مهمی برای فیبروز مرتبط با نفروپاتی دیابتی است که با افزایش بیان TGF-β1 کلیه در شرایط هایپرگلیسمی دائمی بیماران دیابتی و مدلهای حیوانی همراه است. همچنین مشاهده شده که در ادرار بیماران مبتلا به نفروپاتی دیابتی سطح بالایی ازTGF-β وجود دارد که نشان دهنده دخالت TGF-β کلیه در پاتوژنز نفروپاتی دیابتی است (۷). تجزیه و تحلیل همبستگی نشان میدهد که سطوح پلاسمایی TGF-β1 با شدت اختلال عملکرد کلیه در بیماران نفروپاتی دیابتی ارتباط نزدیکی دارد (۵). در مجموع این یافتههای بالینی، پاتولوژیک و اپیدمیولوژیک از بیماران دیابتی و مدلهای حیوانی نشان میدهند که TGF-β1 در بیماریزایی نفروپاتی دیابتی نقش دارد. در سطح سلولی نشان داده شده که سیگنالینگ TGF-β نقش مهمی هم در پیری سلولی و پیری سلولهای بنیادی دارد (۸). به طوریکه مسدودکردن TGF-β با آنتیبادی خنثیکننده، اولیگونوکلئوتیدهای ضد حسگر، مهارکنندهها یا حذف ژنتیکی گیرندهها میتواند باعث کاهش فیبروز کلیه شود (۹). از طرف دیگر، Smad7 (Mothersagainst decapentaplegic homolog 7) به عنوان یک تنظیمکننده مهاری در مسیر سیگنالینگ TGF-β/ SMAD شناخته شده است. SMAD7 میتواند توسط یک مکانیسم وابسته به Smad3 القا شود، که به نوبه خود انتقال سیگنال TGF-β1 را از طریق حلقه بازخورد منفی آن مسدود میکند (۱۰). مطالعات اندکی اهمیت ورزش بر فیبروز ناشی از نفروپاتی دیابتی را گزارش کردهاند (۱4-۱1). مطالعات نشان دادند که فعالیت ورزشی میتواند از پیشرفت نفروپاتی دیابتی در موشهای صحرایی جلوگیری کند (۱3-۱1). همچنین فعالیت ورزشی با شدت متوسط با تقویت Sirt1 (Sirtuin 1) و تنظیم EMT (EPithelial-Mesenchymal Transition) ناشی از TGF-β در دیابت نوع دو از پیشرفت بیماری کلیوی جلوگیری میکند (۱۴). Sirt1 نقش مهمی در هموستاز (Homeostasis) گلوکز و حساسیت به انسولین در کبد، عضلات و بافت چربی دارد (۱۵). بیان این پروتئین با فعالیت آنتی دیابتیک، تنظیم ترشح انسولین، بهبود مسیر سیگنالینگ انسولین و عملکرد میتوکندری، مقاومت به استرس و التهاب ارتباط دارد (۱۵). تمرین با بار فزاینده بر فیبروز کلیوی ناشی از پیری طبیعی، با تنظیم مسیر سیگنالینگ غیر SMAD توسط TGF-β و تحریک فعالشدن اتوفاژی در موشهای مسن فیبروز کلیوی را بهبود میبخشد (۱۶). تمرین تناوبی شدید (High-Intensity Interval Traning, HIIT) ترکیبی از تکرار دورههای فعالیت با شدت بالا (نزدیک به بیشینه یا فوق بیشینه) است که با فعالیت کم شدت تا متوسط و در برخی موارد غیر فعال بودن همراه است (۱۷). HIIT در مقایسه با تمرینات با شدت متوسط علیرغم حجم تمرین کمتر منجر به سازگاریهای مشابه یا حتی بیشتر میشود (۱۸). با وجود مطالعات محدود در رابطه با تاثیر فعالیت ورزشی بر فیبروز کلیوی سالمندان دیابتی و با توجه به اینکه TGF-β هم در سالمندی و هم در بیماری دیابت نقش دارد، بنابراین با مهار مسیرTGF-β1/ SMAD ممکن است فیبروز کلیوی دیابت درمان شود (۱۹). با توجه به اینکه فعالیت ورزشی به عنوان یک راهبرد غیر دارویی در درمان دیابت میباشد، پژوهش حاضر با هدف بررسی تاثیر شش هفته HIIT بر تغییرات بیان ژنهای TGF‐β1 و SMAD7 در موشهای صحرایی دیابتی سالمند مورد ارزیابی قرار گرفت.
روش بررسی
در این پژوهش ۳۶ سر موش صحرایی نر نژاد ویستار به عنوان نمونه پژوهش خریداری شدند. سن موش¬های صحرایی ۲۱ ماه و وزن آنها ۳۴۰ تا ۳۹۰ گرم بود. موشها به روش تصادفی ساده به۳ گروه: کنترل سالم (n=12)، کنترل دیابت (n=12) ، تمرین- دیابت (n=12)، تقسیم شدند. بعد از تایید دیابتی شدن، موشهای صحرایی در قفسهای پلیکربنات مجزا در دمای 22±۲ درجه سانتیگراد و چرخه روشنایی-تاریکی ۱۲ :۱۲ ساعت قرار گرفتند. آشناسازی با پروتکل تمرین تناوبی شدید به مدت یک هفته صورت گرفت. تمام حیوانات دسترسی آزاد به آب و غذا داشتند. القا دیابت موشهای صحرایی: در این پژوهش جهت القا دیابت نوع دو در موشها، محلول استرپتوزوتوسین (STZ) (حلشده در بافر سیترات 0/1 مولار با PH=4/5) به صورت داخل صفاقی فقط یک مرتبه با دوز ۶۰ میلیگرم به ازای هر کیلوگرم از وزن بدن بعد از ۱۵ دقیقه تزریق نیکوتینآمید با دوز ۱۱۰ میلیگرم به ازای هر کیلوگرم از وزن بدن تزریق شد. ۷۲ ساعت بعد از تزریق، قند خون موشها با خونگیری از طریق بریدن نوک دم، بهوسیله گلوکومتر اندازهگیری شد و قند خون ناشتای بالای ۲۰۰ میلیگرم بر دسیلیتر به عنوان دیابت نوع ۲ در نظر گرفته شد. به منظور اطمینان از دیابتی شدن موشهای صحرایی یک هفته بعد قند خون آنها مورد بررسی قرار گرفت و موشهای صحرایی با قند خون ناشتای بالای ۲۰۰ میلیگرم بر دسیلیتر به عنوان مدل دیابت نفروپاتی در نظر گرفته شدند (۲۰،۲۱). آزمون تعیین حداکثر اکسیژن مصرفی: جهت تعیین حداکثر اکسیژن مصرفی از آزمون فزاینده استاندارد Bedford و همکاران (۱۹۷۹) استفاده شد که توسط Leandro و همکاران (۲۰۰۷) برای موشهای صحرایی نژاد ویستار استانداردسازی شده است (۲۲،۲۳). آزمون فزاینده به این صورت بود که موشهای صحرایی بر روی تردمیل با سرعت پنج (متر بر دقیقه) شروع به دویدن کردند و هر سه دقیقه سرعت تردمیل (پنج متر بر دقیقه) افزایش یافت. آزمون تا لحظه رسیدن موشهای صحرایی به واماندگی ادامه داشت. سرعت نهایی موشهای صحرایی به عنوان سرعت بیشینه در زمان رسیدن به حداکثر اکسیژن مصرفی برای محاسبه شدتهای تمرینی موشهای صحرایی استفاده شد.
پروتکل تمرین تناوبی شدید: پروتکل تمرین تناوبی شدید بر مبنای پروتکل تمرین تناوبی شدید رضایی و همکاران طراحی شد با این حال با توجه به تفاوت سنی و دیابتی بودن حیوانات و انجام پایلوت اولیه، پروتکل با توجه به توانایی حیوانات تعدیل شد (۲۴). پروتکل تمرین تناوبی شدید شامل سه قسمت گرمکردن، تمرین (شامل تکرار تناوبها) و سردکردن بود. گرمکردن و سرد کردن موشهای صحرایی با شدت ۴۰ درصد VO2max به مدت ۵ دقیقه دویدن بر روی نوارگردان انجام شد. تمرین تناوبی شامل ترکیبی از تکرار¬های تناوب با شدت بالا و پایین بود. تکرار تناوب با شدت بالا شامل ۲ دقیقه دویدن با شدت ۶۰ درصد VO2max در هفته اول، ۷۰ درصد VO2max در هفته دوم، ۸۰ درصد VO2max در هفته سوم و ۹۰ درصد VO2max از ابتدای هفته چهارم تا پایان هفته ششم بود. تکرار تناوب با شدت پایین در تمام هفتهها شامل ۲ دقیقه دویدن با شدت ۴۰ درصد VO2max بود. تمرین تناوبی به گونه¬ای بود که پس از گرم کردن، موشهای صحرایی ابتدا تناوب با شدت بالا و سپس تناوب با شدت پایین را اجرا کردند. پس از انجام آخرین تکرار تناوب با شدت بالا، موشهای صحرایی به مدت ۵ دقیقه دویدن با شدت40 درصد VO2max سردکردن را انجام دادند. تعداد تکرار تناوب با شدت بالا با توجه به هفته تمرینی موشهای صحرایی تعیین شد. به طوریکه در هفته اول 3 تکرار تناوب، هفته دوم ۵ تکرار تناوب، هفته سوم ۷ تکرار تناوب و از ابتدای هفته چهارم تا پایان هفته ششم شامل ۹ تکرار تناوب بود. از این رو زمان کل تمرین شامل تکرار تناوب با شدت بالا، تکرار تناوب با شدت پایین به همراه گرمکردن و سردکردن در هفته اول ۲۰ دقیقه، در هفته دوم ۲۸ دقیقه، هفته سوم ۳۶ دقیقه و از ابتدای هفته چهارم به بعد ۴۴ دقیقه بود. شیب دستگاه در کل پروتکل تمرینی صفر در نظر گرفته شد. (مطابق با جدول ۱). گروههای کنترل سالم و دیابت در این مدت هیچ گونه تمرینی انجام ندادند (24-22).
اندازهگیری میزان بیان ژنهای TGF‐β1 و SMAD7: بیست و چهار ساعت پس از آخرین جلسه تمرینی، موشها با ترکیبی از کتامین (mg/kg۹۰) و زایلازین (mg/kg۱۰) بیهوش شدند (۲۵). بافت کلیه تحت شرایط استریل جدا شد و بلافاصله در نیتروژن مایع (دمای ۱۹۶-) منجمد شد و تا زمان فرا رسیدن انجام کار استخراج RNA در فریزر ۸۰- درجه نگهداری شدند. برای بررسی بیان ژنهای TGF-1 و SMAD7 در بافت کلیه موشهای صحرایی با استفاده از کیت ستونی استخراج RNA (FavorPreP™ Tissue Total RNA Mini Kit) ساخت کشور هنگکنگ طبق دستورالعمل (FATRK 001)، کل محتویات RNA سلول (total RNA) استخراج شد. بافت با استفاده از یک میلیمول RNAx هموژن شد سپس با اضافه کردن ترکیبی از RB بافر و بتامرکاپتواتانول (MercaPtoethanol -) نمونه لیز شده و با دستگاه همگن کننده بافت هموژن گردید. در مرحله بعد نمونه روی فیلتر سفید گذاشته شد و به مدت ۳۰ دقیقه سانتریفیوژ گردید. ماده زیر توری سفید بدون رسوب ایجاد شده در آن به میکروتیوب دیگری منتقل شد. RNA استخراج شده با ۴۵۰ میکرولیتر اتانول ۷۰% شستشو و خشک گردید و سپس 1/500 میکرولیتر آب اضافه شد و ۳۰ دقیقه سانتریفیوژ گردید. در مرحله بعد ۷۵۰ میکرولیتر آب استریل اضافه و ۳۰ دقیقه سانتریفیوژ گردید. این مرحله دو مرتبه تکرار شد. سپس RNAase free water اضافه شد و ۳۰ دقیقه با دور بالا سانتریفیوژ شد. کمیت و کیفیت RNA استخراج شده با استفاده از اسپکتروفتومتری نانو دراپ با نسبت چگالی نوری 280/260 نانومتر و الکتروفورز بر روی ژل آگارز (1%) ارزیابی شد (دستگاه پیکو دراپ شرکت سیگما ساخت امریکا). سنتز cDNA طبق دستورالعمل موجود در کیت فرمنتاز (K1621) تهیه گردید. هنگام تهیه cDNAاز نمونه تخلیص شده پس از قرائت جذب، حجمی شامل ۱۰۰۰ نانوگرم RNA برداشته شد، سپس 0/5 میکرولیتر رندوم هگزامر (Random Hexamers)، 0/5 میکرولیتر پرایمر الیگو دی تی (Oligo dT) به آن افزوده و تا حجم ۱۲ میکرولیتر آب DEPC اضافه گردید و به دمای 65 درجه به مدت 5 دقیقه منتقل شد. سپس به مدت 2 دقیقه بر روی یخ قرار گرفت تا ساختارهای ثانویه RNA از هم باز شوند. در مرحله بعد ۴ میکرولیتر 5X Reaction Buffer، ۲ میکرولیتر dNTP، ۱ میکرولیتر RiboLock RNase Inhibitorو ۱ میکرولیتر RevertAid RT به ترکیب قبل اضافه گردید. سپس ترکیب ابتدا به مدت 5 دقیقه در دمای 25 درجه قرار گرفت. بعد از آن به مدت 60 دقیقه در دمای 42 درجه قرار گرفت. در آخر به منظور از کار افتادن آنزیم RT، تیوبهای واکنش به مدت 5 دقیقه در دمای 70 درجه سانتیگراد قرار گرفت. سپس cDNA آماده شده جهت انجام RT- PCR مورد استفاده قرار گرفت. تمام پرایمرها توسط نرمفزار Allele IDv7.8 طراحی شد و از ژن خانهبان (HousekeePing) B2m به عنوان کنترل داخلی واکنشهای qPCR استفاده شد. تمام پرایمرها به صورت اتصال اگزون- اگزون طراحی شدند. جهت اطمینان از عدم تکثیر DNA ژنومی از 25 نانوگرم cDNA و 25 نانوگرم RNA در تیوبهای جداگانه از واکنش PCR و بهکارگیری از ژل آگاروز 5/1% استفاده شد. همه نمونهها برای شرایط qPCR آماده شدند و در دمای 94 درجه سانتیگراد به مدت 10 دقیقه به دنبال 40 سیکل 15 ثانیهای در دمای 94 درجه سانتیگراد، 60 ثانیه در دمای 58 درجه سانتیگراد و سپس 7 دقیقه در دمای 72 درجه سانتیگراد قرار گرفتند (۲۶). برای کمی سازی مقادیر بیان ژن از فرمول ct-۲ (۲به توان منفی ct) استفاده شد و مقادیر Fold change محاسبه شد (۲۷). تمام آزمایشات RT-PCR در سه تکرار انجام شد. توالی پرایمرهای پژوهش در جدول ۲ ارائه شده است.
تجزیه و تحلیل آماری
برای بررسی طبیعی بودن توزیع دادهها از آزمون شاپیرو-ویلک و جهت بررسی تفاوت معناداری بین گروهها از آزمون تحلیل واریانس آنوا یک طرفه و آزمون تعقیبی توکی استفاده شد. تجزیه و تحلیلهای آماری با استفاده از نرمافزارSPSS version 16 در سطح معناداری P<0/05 انجام شد.
ملاحظات اخلاقی
پروپوزال این پژوهش، مورد تایید کمیته اخلاق در پژوهش دانشگاه علوم پزشکی شیراز (IR.SUMS.REHAB.REC.1400.027) قرار گرفته است.
نتایج
نتایج تحلیل واریانس آنوای یک طرفه نشان داد که بهطور کلی بین وزن بدن موشهای صحرایی در گروههای مختلف تفاوت معناداری وجود دارد (P= 0/000، F=26/47). القای دیابت منجر به کاهش وزن بدن موشهای صحرایی شد (P=0/000)، همچنین بین وزن موشهای صحرایی در گروههای دیابت تمرین و کنترل دیابت تفاوت معناداری وجود نداشت (P=0/13). نتایج آزمون تعقیبی توکی در جدول ۳ نشان داده شده است. نتایج تحلیل واریانس آنوای یک طرفه تفاوت معناداری را در بیان ژن TGF-β1 بافت کلیه در بین گروه¬های مورد مطالعه نشان داد (P=0/000، F= 34/39) (جدول 4). نتایج آزمون تعقیبی توکی نشان داد که گروه تمرین- دیابت کاهش معناداری در بیان ژن TGF-β1 بافت کلیه نسبت به گروه دیابت داشت (P=0/001). همچنین القا دیابت منجر به افزایش بیان ژن TGF-β1 (P=0/000) در بافت کلیه موش¬های صحرایی سالمند نسبت به گروه کنترل سالم شد (0/001=P) (شکل ۱). علاوه بر این تفاوت معناداری در بیان ژن SMAD7 در بافت کلیه در بین گروههای مورد مطالعه وجود داشت (P=0/000، F= 17/84) (جدول 5). نتایج آزمون تعقیبی توکی نشان داد که گروه تمرین- دیابت نسبت به گروه کنترل- دیابت منجر به افزایش معناداری در بیان ژن SMAD7 در بافت کلیه شد (0/008=P). همچنین بین گروه دیابت و کنترل سالم در بیان ژن SMAD7 کاهش معناداری مشاهده شد (0/001=P) (شکل۲).
جدول ۱: پروتکل تمرینی مورد استفاده در پژوهش
جدول ۲: توالی پرایمرهای استفاده شده در این پژوهش
جدول ۳: میانگین ± انحراف معیار وزن بدن موشهای صحرایی (گرم) و نتایج آزمون تعقیبی توکی در گروههای مختلف
علامت * نشان دهنده تفاوت معنادار است
جدول4: نتایج تحلیل واریانس آنوای یک طرفه TGF-1
علامت * نشان دهنده تفاوت معنادار است
شکل۱: تغییرات بیان mRNA TGF-β1 در گروه¬های کنترل سالم (Control)، کنترل دیابت (Diabetes) و دیابت- تمرین (Diabetes+ HIIT).
علامت * نشان دهنده تفاوت معنادار است
جدول5: نتایج تحلیل واریانس آنوای یک طرفه SMAD7
(Smad7 (Mothers against decapentaplegic homolog 7 علامت * نشان دهنده تفاوت معنادار است
شکل۲: تغییرات بیان mRNA SMAD7 در گروههای کنترل سالم (Control)، کنترل- دیابت (Diabetes) و دیابت- تمرین (Diabetes+ HIIT). علامت * نشان دهنده تفاوت معنادار است.
بحث
با تغییر سبک زندگی و روند پیری جمعیت، بروز دیابت نوع دو سال به سال در حال افزایش است. شیوع دیابت یک مشکل رو به رشد بهداشت عمومی جهانی است (۲۸). نفروپاتی دیابتی یک عارضه میکروواسکولار (Microvascular) جدی در بیماران دیابتی است که با گلومرولواسکلروزیس (Glomerulosclerosis) ناشی از متابولیسم غیرطبیعی گلوکز مشخص میشود و همچنین علت اصلی مرگ در نفروپاتی دیابتی است (۲۹). پیشرفت این بیماری به تدریج میتواند منجر به کاهش عملکرد کلیوی، ادم سیستمیک و حتی نارسایی کلیه در مرحله پایانی تبدیل شود که یکی از علل شایع مرگ در بیماران دیابتی است (۳۰،۳۱). گرچه ورزش آثار مفید مختلفی در دیابت نوع دو و چاقی دارد، اما اثرات کلیوی ورزش، بهخصوص HIIT و مکانیسمهای زیربنایی مسئول هنوز به طور کامل مشخص نشده است. پژوهش حاضر به منظور بررسی تاثیر شش هفته تمرین تناوبی شدید بر بیان ژن TGF-β1 و SMAD7 در بافت کلیه در موشهای صحرایی سالمند دیابتی صورت گرفت. نتایج مطالعه نشان داد که تمرین تناوبی شدید آثار محافظتی در برابر نفروپاتی دارد. در پژوهش حاضر برای شناخت تغییرات فیزیولوژیکی بافت کلیه در پاسخ به تمرینات تناوبی شدید، TGF-β1 مورد بررسی قرار گرفت. نتایج پژوهش نشان داد که HIIT منجر به کاهش معناداری در بیان ژن TGF-β1 در بافت کلیه میشود. همسو با این نتایج Ren و همکاران (۲۰۱۷) نشان دادند که فعالیت ورزشی با شدت متوسط با تقویت Sirt1 و تنظیم EMT ناشی از TGF-β در دیابت نوع دو از پیشرفت بیماری کلیوی جلوگیری میکند (۱۴). به علاوه BAO و همکاران (۲۰۱۹) تاثیر تمرین با بار فزاینده بر فیبروز کلیوی ناشی از پیری طبیعی و مکانیسمهای آن را مورد مطالعه قرار دادند و نشان دادند که تمرین فزاینده با تنظیم مسیر سیگنالینگTGF‐β1/ TAK1/ MKK3/ P38MAPK (مسیرسیگنالینگ غیر SMAD) و تحریک فعالشدن اتوفاژی در موشهای مسن فیبروز کلیوی را بهبود میبخشد (۱۶). با وجود این برخی تحقیقات افزایش سطح TGF-β را بر اثر تمرین در بافتهای دیگر گزارش کردهاند (۳۲،۳۳). مقدم و همکاران (۱۳۹۶) اثر حفاظتی تمرین هوازی بر سرطان پستان بهواسطه پروتئین TGF-β در موشهای ماده را مورد ارزیابی قرار دادند. نتایج آنها نشان داد که ده هفته تمرین استقامتی هوازی منجر به افزایش معنادار میانگین مقادیر پروتئین TGF-β در بافت تومور نسبت به گروه کنترل شد (۳۳). احتمالاً دلیل افزایش TGF-β متفاوت بودن نوع تمرین باشد. در پژوهش دیگری نشان داده شد که هشت هفته تمرین هوازی منجر به افزایش مقدار TGF-β در بافت قلب موشهای صحرایی سالمند میشود (۳۲). این افزایش شاید به دلیل کافی نبودن شدت در برنامه تمرینی مطالعه فوق باشد. نشان داده شده که هایپرگلیسمی حاد و مزمن بیان TGF-β1 را القا میکند. TGF-β1 یک سایتوکاین اصلی در چاقی و مقاومت به انسولین است و همچنین با داشتن خواص فیبروژنیک نقش محوری در ایجاد نفروپاتی دیابتی دارد (۳۴). عوامل موثر بر بیان TGF-β1 بیشمار هستند (۳۵). هایپرگلیسمی یکی از عوامل اصلی بیان TGF-β1 است و بیماران دیابتی سطح TGF-β1 بالاتری نسبت به افراد سالم دارند. تصور میشود که مسیر دی آسیل گلیسرول پروتئین کیناز C، مسیر گلوکوزآمین و احتمالاً مسیر پلیول، نقش مهمی در تولید بیشتر TGF-β1در دیابت دارند (۳۶). یافتههای محققان حاکی از این است که گلوکز واسطه اصلی بیان TGF-β1است (۳۷). هایپرگلیسمی که معمولاً در بیماران مبتلا به دیابت نوع دو مشاهده میشود با بیش تنظیمی انتقالدهنده گلوکز-۱ (GLUT1) همراه است که منجر به بیان بیش از حد TGF-β توسط سلولهای توبولی مزانژیال یا سلولهای کلیوی میشود (۳۸،۳۹). علاوه بر این افزایش فشار داخل گلومرولی، کشش سلول مزانژیال، فعال شدن سیستم رنین-آنژیوتانسین، گونههای فعال اکسیژن (ROS) و محصولات نهایی گلیکاسیون پیشرفته (AGEs) احتمالاً باعث تولید TGF-β در کلیه یا در سلولهای مزانژیال یا توبولی میشود (۳۹،۴۰). TGF-β از طریق مسیرهای اتوکرین و پاراکرین عمل کرده و گیرندههای سیگنالینگ و سنتز ECM آن را تحریک میکنند (۳۹،۴۰). شواهد بالینی نشان میدهد که پاسخ ناشی از TGF-β شامل یک مسیر نهایی است که منجر به توسعه گلومرولو اسکلروز و فیبروز بینابینی در مرحله پایانی بیماری کلیوی (ESRD) میشود (۴۱). با توجه به مکانسیم فوق کاهش میزان هایپرگلیسمی و مقاومت به انسولین در گروه HIIT شاید منجر به کاهش بیان TGF-β1 در بافت کلیه شود که بیانگر این است که تمرین تناوبی شدید میتواند اثرات مخرب مسیر سیگنالینگ TGF-β1 را کاهش داده و از شدت آسیب به بافت کلیه در بیماران نفروپاتی دیابتی جلوگیری کند. یکی دیگر از نتایج پژوهش حاضر افزایش معنادار بیان ژن SMAD7 در بافت کلیه در پاسخ به HIIT بود. مطالعات اندکی تاثیر تمرینات ورزشی را بر بیان mRNA SMAD7 در بافت کلیه در بیماری دیابت را بررسی کردهاند. در این راستا Hou و همکاران (۲۰۲۱) نشان دادند که تمرینات شنا به مدت ۱۲ هفته بیان mRNA SMAD7 میوکارد در موش-های صحرایی دیابتی را افزایش میدهد و منجر به کاهش معناداری در قند خون و بیان mRNA TGF-β1 میشود (۴۲). همچنین Wang و Yuan (۲۰۱۹) گزارش کردند که مدل تمرین تناوبی سرعتی (SIT) میتواند منجر به تنظیم مثبت SMAD7 در بافت کلیه موشهای صحرایی دیابتی شود (۴۳). اجزای اساسی سیگنالینگ TGF-β1 شامل یک کمپلکس گیرنده با گیرندههای مرتبط با غشا نوع I و نوع II و پروتئینهای SMAD است که در عوامل رونویسی پایین دستی نقش دارند (۴۴،۴۵). SMAD7به عنوان یک تنظیم کننده مهاری در مسیر سیگنالینگ TGF-β/ SMAD میتواند توسط یک مکانیسم وابسته به Smad3 القا شود، که به نوبه خود انتقال سیگنال TGF-β1 را از طریق حلقه بازخورد منفی آن مسدود میکند. SMAD7 با مسدود کردن دسترسی R-Smadها به TβRI و یا با افزایش تخریب کمپلکسهای گیرنده، فسفوریلاسیون R-Smad را مهار میکنند (۱۰). در مقابل شکل فعال TGF-β، شکل نهفته TGF-β1 میتواند با بیش تنظیم SMAD7 در موشهای ترانسژنیک از کلیهها در برابر فیبروز و التهاب محافظت کند (۱۰). گزارشات نشان میدهد که القای SMAD7 گذار اپیتلیال مزانشیمی توبولی و ایجاد آسیبهای فیبروتیک را بلاک میکند (۴۶). نقش عملکردی SMAD7 بیشتر با یافتههایی مشخص میشود که حذف SMAD7 باعث تسریع فیبروژنز کلیه در نفروپاتی انسدادی، نفروپاتی دیابتی و همچنین نفروپاتی فشار خون میشود (۴۹-۴۷)، که SMAD7 را به عنوان یک عامل درمانی برای درمان بیماریهای مزمن کلیوی نشان میدهد (49-52،۴7). SMAD7 سیگنالهای TGF-β/ SMAD را از طریق دو مکانیسم ممکن بهطور منفی تنظیم میکند. نخست اینکه SMAD7 به گیرنده TGF-β1 متصل میشود و در نتیجه از جذب و فسفوریلاسیون Smad2 و Smad3 جلوگیری میکند و دیگر اینکه SMAD7به عنوان یک گیرنده عمل میکند (۵۳). پروتئین آداپتوری که لیگازهای یوبیکوئیتین مانند Smurf2 را به مجموعه گیرنده TGF-β جذب میکند تا تخریب آن را از طریق مسیرهای تخریب پروتئازوم- یوبیکوئیتین تقویت کند (۵6-۵4). با توجه به مکانسیم فوق، تمرین تناوبی شدید با کاهش بیان TGF-β1 در بافت کلیه و تاثیرش بر افزایش بیان SMAD7 میتواند منجر به اتصال SMAD7 به گیرنده TGF-β1 شده و فسفریلاسیون R-Smad را مهار کند و یا با افزایش تخریب کمپلکسهای گیرنده از آسیبهای فیبروتیک جلوگیری کند. بنابراین با توجه به نتایج پژوهش حاضر شاید HIIT بهتواند به عنوان یک روش درمانی غیر دارویی در جلوگیری از پیشرفت نفروپاتی دیابتی، از طریق مسدود کردن فیبروز کلیه با واسطه TGF-β1 و تنظیم مثبت SMAD7 عمل کند که ممکن است اساس درمان بیماران نفروپاتی دیابتی را در آینده فراهم کند. از آنجایی که AGEs یک واسطه ضروری در عوارض دیابت هستند، این محصولات قادر به فعالکردن Smad 2/3 به طور مستقیم و مستقل از TGF-β هستند و توسط گیرنده AGEs از طریق مسیر تداخلی وابسته به MAP کیناز ERK/P38 عمل میکنند. با توجه به اینکه مطالعه حاضر تاثیر HIIT بر مسیر TGF-β /Smad را مورد بررسی قرار داده لذا لازم به ذکر است که در پژوهشهای آینده نیاز است تا تاثیر HIIT بر مسیر ERK/P38 در موشهای دیابتی سالمند مورد بررسی قرار گیرد. از محدودیتهای این پژوهش میتوان به عدم اندازهگیری گیرنده TGF-β1 و فاکتورهای درگیر در مسیر غیروابسته به SMAD اشاره کرد.
نتیجهگیری
بهطور کلی مطالعه حاضر نشان داد که القای دیابت منجر به افزایش معناداری در بیان mRNA TGF-β1 و کاهش معناداری در بیان mRNA SMAD7 میشود و شش هفته تمرین تناوبی شدید به ترتیب منجر به کاهش و افزایش معنادار در mRNA TGF-β1 و mRNA SMAD7 در بافت کلیه موش¬های صحرایی دیابتی میشود و در نتیجه میتواند در کلیهها اثرات محافظتی در برابر نفروپاتی را اعمال کند.
سپاسگزاری
مقاله حاضر برگرفته از رساله دکتری دانشجویی دانشگاه شیراز میباشد، لذا نویسندگان مراتب تقدیر و تشکر خود را از کلیه افرادی که در اجرای این پژوهش ایفای نقش کردند، اعلام میدارند.
حامی مالی: ندارد.
تعارض در منافع: وجود ندارد.
References:
1- Shlisky J, Bloom DE, Beaudreault AR, Tucker KL, Keller HH, Freund-Levi Y, et al. Nutritional Considerations for Healthy Aging and Reduction in Age-Related Chronic Disease. Adv in Nutr 2017; 8(1): 17-26.
2- Chang AS, Hathaway CK, Smithies O, Kakoki M. Transforming Growth Factor-Β1 and Diabetic Nephropathy. American J Physiology-Renal Physiology 2016; 310(8): 689-96.
3- Association AD. Standards of Medical Care in Diabetes-2014. Diabetes Care 2014; 37: 14-80.
4- Dekkers CC, Gansevoort RT, HeersPink HJ. New Diabetes Therapies and Diabetic Kidney Disease Progression: The Role of SGLT-2 Inhibitors. Current Diabetes Reports 2018; 18(5): 1-12.
5- Ibrahim S, Rashed L. Estimation of Transforming Growth Factor-Beta 1 as a Marker of Renal Injury in Type II Diabetes Mellitus. Saudi Med J 2007; 28(4): 519-23.
6- Jakuš V, SaPák M, Kostolanská J. Circulating TGF-Β1, Glycation, and Oxidation in Children with Diabetes Mellitus Type 1. ExP Diabetes Res 2012; 2012: 510902.
7- Shaker YM, Soliman HA, Ezzat E, Hussein NS, Ashour E, Donia A, et al. Serum and Urinary Transforming Growth Factor Beta 1 as Biochemical Markers in Diabetic Nephropathy Patients. Beni-Suef University J Basic and APPlied Sciences 2014; 3(1): 16-23.
8- Tominaga K, Suzuki HI. TGF-Β Signaling in Cellular Senescence and Aging-Related Pathology. International J Molecular Sciences 2019; 20(20): 5002.
9- Hills CE, Squires PE. The Role of TGF-Β and Epithelial-To Mesenchymal Transition in Diabetic Nephropathy. Cytokine Growth Factor Rev 2011; 22(3): 131-9.
10- Meng X-M, Tang PM-K, Li J, Lan HY. TGF-Β/Smad Signaling in Renal Fibrosis. Front Physiol 2015; 6: 82.
11- Boor P, Celec P, Behuliak M, Grančič P, Kebis A, Kukan M, et al. Regular Moderate Exercise Reduces Advanced Glycation and Ameliorates Early Diabetic Nephropathy in Obese Zucker Rats. Metabolism 2009; 58(11): 1669-77.
12- Chiasera JM, Ward-Cook KM, McCune SA, Wardlaw GM. Effect of Aerobic Training on Diabetic Nephropathy in a Rat Model of Type 2 Diabetes Mellitus. Ann Clin Lab Sci 2000; 30(4): 346-53.
13- Ito D, Cao P, Kakihana T, Sato E, Suda C, Muroya Y, et al. Chronic Running Exercise Alleviates Early Progression of Nephropathy with Upregulation of Nitric Oxide Synthases and Suppression of Glycation in Zucker Diabetic Rats. PloS one 2015; 10(9): e0138037.
14- Ren L, Sen U, PushPakumar S. Exercise Training Reduces TGF‐Β Mediated Epithelial Mesenchymal Transition in Diabetic Kidney. The FASEB Journal 2017; 31(S1): 1086-5.
15- Cantó C, Auwerx J. PGC-1alpha, SIRT1 and AMPK, an Energy Sensing Network that Controls Energy Expenditure. Curr Opin Lipidol 2009; 20(2): 98-105.
16- Bao C, Yang Z, Cai Q, Li Q, Li H, Shu B. Incremental Load Training Improves Renal Fibrosis by Regulating the TGF‑Β1/TAK1/MKK3/P38MAPK Signaling Pathway and Inducing the Activation of Autophagy in Aged Mice. International J Molecular Med 2019; 44(5): 1677-86.
17- Laursen PB, Jenkins DG. The Scientific Basis for High-Intensity Interval Training. Sports Med 2002; 32(1): 53-73.
18- Winding KM, Munch GW, IePsen UW, Van Hall G, Pedersen BK, Mortensen SP. The Effect on Glycaemic Control of Low Volume High Intensity Interval Training Versus Endurance Training in Individuals with Type 2 Diabetes. Diabetes, Obesity and Metabolism 2018; 20(5): 1131-9.
19- Wu W, Huang XR, You Y, Xue L, Wang X-J, Meng X, et al. Latent TGF-Β1 Protects Against Diabetic Kidney Disease Via Arkadia/Smad7 Signaling. Int J Biol Sci 2021; 17(13): 3583-94.
20- Pierre W, Gildas AJH, Ulrich MC, Modeste WN, Albert K. Hypoglycemic and Hypolipidemic Effects of Bersama Engleriana Leaves in Nicotinamide/Streptozotocin-Induced Type 2 Diabetic Rats. BMC Complement Altern Med 2012; 12(1): 264.
21- Maheshwari R, Balaraman R, Sen AK, Shukla D, Seth A. Effect of Concomitant Administration of Coenzyme Q10 with Sitagliptin on Experimentally Induced Diabetic Nephropathy in Rats. Ren fail 2017; 39(1): 130-9.
22- Bedford TG, TiPton CM, Wilson NC, OPPliger RA, Gisolfi CV. Maximum Oxygen Consumption of Rats and Its Changes with Various Experimental Procedures. J Appl Physiol Respir Environ Exerc Physiol 1979; 47(6):1278-83.
23- Leandro C G, Levada AC, Hirabara SM, Manhães-de-Castro R, De-Castro CB, Curi R, et al. A Program of Moderate Physical Training for Wistar Rats Based on Maximal Oxygen Consumption. J Strength and Cond Res 2007; 21(3): 751-6.
24- Rezaei R, Nasoohi S, HaghParast A, Khodagholi F, Bigdeli MR, Nourshahi M. High Intensity Exercise Preconditioning Provides Differential Protection Against Brain Injury Following Experimental Stroke. Life Sciences 2018; 207: 30-5.
25- Kumar AH, Clover AJ. Intraperitoneal Co-Administration of Low Dose Urethane with Xylazine and Ketamine for Extended Duration of Surgical Anesthesia in Rats. Lab Anim Res 2015; 31(4): 174-9.
26- Vafaei H, Kavari G, Izadi HR, Dorahi ZZ, DianatPour M, DaneshParvar A, et al. Wi-Fi (2.4 Ghz) Affects Antioxidant Capacity, DNA Repair Genes Expression and, Apoptosis in Pregnant Mouse Placenta. Iran J Basic Med Sci 2020; 23(6): 833-40.
27- Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real Time Quantitative PCR and the 2− ΔΔCT Method. Methods 2001; 25(4): 402-8.
28- Sugiyama T, Goryoda S, Inoue K, Sugiyama-Ihana N, Nishi N. Construction of a Simulation Model and Evaluation of the Effect of Potential Interventions on the Incidence of Diabetes and Initiation of Dialysis Due to Diabetic Nephropathy in Japan. BMC Health Serv Res 2017; 17(1): 833.
29- Sharma D, Bhattacharya P, Kalia K, Tiwari V. Diabetic Nephropathy: New Insights into Established Therapeutic Paradigms and Novel Molecular Targets. Diabetes Res Clin Pract 2017; 128: 91-108.
30- Warren AM, Knudsen ST, CooPer ME. Diabetic Nephropathy: An Insight Into Molecular Mechanisms And Emerging Therapies. Expert Opin Ther Targets 2019; 23(7): 579-91.
31- Zürbig P, Mischak H, Menne J, Haller H. CKD273 Enables Efficient Prediction of Diabetic Nephropathy in Nonalbuminuric Patients. Diabetes Care 2019; 42(1): e4-e5.
32- Goodarzi F, Abednatanzi H, Nikbakht H, Ebrahim Kh, Ghazaliyan F. Effects of Eight Weeks Aerobic Exercise on the Signaling Pathway of Cardiac Fibrosis in Elderly Rats. Journal of Knowledge & Health in Basic Medical Sciences 2020; 14(4): 48-53.[Persian]
33- Moghadam V, Piri M, Azarbayjani MA, Matinhomaee H. The Protective Effect of Aerobic Exercise on Breast Cancer by Tgfβ Protein and Smad-3 and MMP2 Gene in Female Mice. SJKU 2017; 22(3): 60-73. [Persian]
34- Wang M, Zhang J, SPinetti G, Jiang L-Q, Monticone R, Zhao D, et al. Angiotensin II Activates Matrix Metalloproteinase Type II and Mimics Age-Associated Carotid Arterial Remodeling in Young Rats. Am J Pathol 2005; 167(5):1429-42.
35- Tzavlaki K, Moustakas A. TGF-β Signaling. Biomolecules 2020; 10(3): 487.
36- Rossert J, Terraz-Durasnel C, Brideau G. Growth Factors, Cytokines, and Renal Fibrosis During the Course of Diabetic Nephropathy. Diabetes Metab 2000; 26 Suppl 4: 16-24.
37- HeydarPour F, Sajadimajd S, Mirzarazi E, HaratiPour P, Joshi T, Farzaei MH, et al. Involvement of TGF-Β and Autophagy Pathways in Pathogenesis of Diabetes: A Comprehensive Review on Biological and Pharmacological Insights. Front Pharmacol 2020; 11: 498758.
38- El-Sherbini SM, Shahen SM, Mosaad YM, Abdelgawad MS, Talaat RM. Gene Polymorphism of Transforming Growth Factor-Β1 in Egyptian Patients with Type 2 Diabetes and Diabetic Nephropathy. Acta Biochim BioPhys Sin 2013; 45(4): 330-8.
39- Qian Y, Feldman E, Pennathur S, Kretzler M, Brosius III FC. From Fibrosis to Sclerosis: Mechanisms of Glomerulosclerosis in Diabetic Nephropathy. Diabetes 2008; 57(6): 1439-45.
40- Sharma K, Ziyadeh FN. Hyperglycemia and Diabetic Kidney Disease: The Case for Transforming Growth Factor–Β as a Key Mediator. Diabetes 1995; 44(10):1139-46.
41- Border WA, Noble NA, Yamamoto T, HarPer JR, Yamaguchi Y, Pierschbacher MD, et al. Natural Inhibitor of Transforming Growth Factor-Β Protects Against Scarring in Experimental Kidney Disease. Nature 1992; 360(6402): 361-4.
42- Hou G, Liu Q, Xi X, Liu H. Effects of Swimming And Epigallocatechin Gallate on Interstitial Proteins Expression of Myocardium From Type 2 Diabetic Rats. Wei Sheng yan jiu 2021; 50(1): 86-92.
43- Wang S-Q, Li D, Yuan Y. Long-Term Moderate Intensity Exercise Alleviates Myocardial Fibrosis in Type 2 Diabetic Rats Via Inhibitions of Oxidative Stress and TGF-Β1/Smad Pathway. J Physiol Sci 2019; 69(6): 861-73.
44- Xie F, Ling L, van Dam H, Zhou F, Zhang L. TGF-Β Signaling in Cancer Metastasis. Acta Biochim Biophys Sin (Shanghai) 2018; 50(1):121-32.
45- Zhang YE. Mechanistic Insight Into Contextual TGF-Β Signaling. Curr Opin Cell Biol 2018; 51: 1-7.
46- Saika S, Ikeda K, Yamanaka O, Sato M, Muragaki Y, Ohnishi Y, et al. Transient Adenoviral Gene Transfer of Smad7 Prevents Injury Induced Epithelial Mesenchymal Transition of Lens Epithelium in Mice. Lab invest 2004; 84(10):1259-70.
47- Chen HY, Huang XR, Wang W, Li JH, Heuchel RL, Chung AC, et al. The Protective Role of Smad7 in Diabetic Kidney Disease: Mechanism and Therapeutic Potential. Diabetes 2011; 60(2): 590-601.
48- Chung AC, Huang XR, Zhou L, Heuchel R, Lai KN, Lan HY. Disruption of the Smad7 Gene Promotes Renal Fibrosis and Inflammation in Unilateral Ureteral Obstruction (UUO) in Mice. Nephrol Dial Transplant 2009; 24(5): 1443-54.
49- Liu G-X, Li Y-Q, Huang XR, Wei LH, Zhang Y, Feng M, et al. Smad7 Inhibits Angii-Mediated Hypertensive Nephropathy in a Mouse Model of Hypertension. Clin Sci 2014; 127(3): 195-208.
50- Ka S, Yeh Y, Huang X, Chao T, Hung Y, Yu C, et al. Kidney-Targeting Smad7 Gene Transfer Inhibits Renal TGF-Β/MAD Homologue (SMAD) and Nuclear Factor Κb (NF-Κb) Signalling Pathways, and Improves Diabetic Nephropathy in Mice. Diabetologia 2012; 55(2): 509-19.
51- Ka S-M, Huang X-R, Lan H-Y, Tsai P-Y, Yang S-M, Shui H-A, et al. Smad7 Gene Therapy Ameliorates an Autoimmune Crescentic Glomerulonephritis in Mice. J Am Soc NePhrol 2007; 18(6): 1777-88.
52- Lan HY, Mu W, Tomita N, Huang XR, Li JH, Zhu H-J, et al. Inhibition of Renal Fibrosis by Gene Transfer of Inducible Smad7 Using Ultrasound-Microbubble System in Rat UUO Model. J Am Soc NePhrol 2003; 14(6): 1535-48.
53- Zhu H-J, Iaria J, Sizeland AM. Smad7 Differentially Regulates Transforming Growth Factor Β-Mediated Signaling Pathways. J Biol Chem 1999; 274(45): 32258-64.
54- Ebisawa T, Fukuchi M, Murakami G, Chiba T, Tanaka K, Imamura T, et al. Smurf1 Interacts with Transforming Growth Factor-Β Type I Receptor through Smad7 and Induces Receptor Degradation. J Biol Chem 2001; 276(16): 12477-80.
55- Kavsak P, Rasmussen RK, Causing CG, Bonni S, Zhu H, Thomsen GH, et al. Smad7 Binds to Smurf2 to Form an E3 Ubiquitin Ligase that Targets the Tgfβ Receptor for Degradation. Mol cell 2000; 6(6): 1365-75.
56- Liu F-Y, Li X-Z, Peng Y-M, Liu H, Liu Y-H. Arkadia-Smad7-Mediated Positive Regulation of TGF-Β Signaling in a Rat Model of Tubulointerstitial Fibrosis. Am J Nephrol 2007; 27(2): 176-83.
نوع مطالعه: پژوهشي |
موضوع مقاله:
فیزیولوژی ورزش
دریافت: 1401/6/6 | پذیرش: 1401/8/22 | انتشار: 1401/12/15
دریافت: 1401/6/6 | پذیرش: 1401/8/22 | انتشار: 1401/12/15
ارسال پیام به نویسنده مسئول
| بازنشر اطلاعات | |
 |
این مقاله تحت شرایط Creative Commons Attribution-NonCommercial 4.0 International License قابل بازنشر است. |
